
Enrico Di Luise
Tenente dei Carabinieri, ufficiale addetto alla Sezione di Biologia Molecolare presso il Reparto Carabinieri Investigazioni Scientifiche (R.I.S.) di Messina. Laureato in Scienze e Tecnologie Alimentari all’Università di Catania.

Paola Magni
Laureata in Scienze Naturali presso la Facoltà di Scienze Matematiche, Fisiche e Naturali dell'Università di Torino. Lavora presso la Direzione della sezione di Entomologia Forense del Forensic Lab dell'ICAA (International Crime Analysis Association).

Luigi Saravo
Maggiore dei Carabinieri, ufficiale addetto al Reparto Carabinieri Investigazioni Scientifiche (R.I.S.) di Roma. Laureato in Scienze Biologiche all'Università di Napoli “Federico II”.
1. Prefazione
Per molti anni gli insetti che popolavano la superficie e gli orifizi dei cadaveri sono stati considerati solo una presenza poco gradevole del lavoro dei medici legali e delle Forze dell’Ordine coinvolte nelle indagini inerenti a crimini violenti. Ma come gli intriganti romanzi di Deawer e della Cornwell ci hanno insegnato, gli insetti costituiscono dei silenziosi custodi di preziosi informazioni sull’accadimento di fatti a volte inspiegabili. Tuttavia mentre le varie branche delle scienze criminalistiche (la biologia, la balistica, la chimica, ecc.) hanno avuto nel tempo una crescita esponenziale, affinando tecniche e metodologie, lo studio degli insetti associati alla scena criminis è stato, nel nostro Paese per molto tempo sottovalutato o addirittura ignorato.
L’Entomologia Forense è un ramo della zoologia dedicata allo studio degli insetti e di altri artropodi, dal cui esame è possibile ricavare elementi decisivi per la persecuzione di un reato. Negli ultimi anni tale disciplina ha visto crescere l’interesse da parte degli scienziati forensi, dei Medici Legali e delle Forze dell’Ordine, soprattutto in Europa e negli USA, dove esistono figure professionali specialistiche che affiancano la magistratura inquirente nella risoluzione di casi di morti sospette ed altri reati accomunati dalla presenza di insetti. Purtroppo in Italia solo da pochi anni si è sviluppato l’interesse verso la materia e ancora oggi gli entomologi forensi non lavorano sistematicamente accanto agli organi di Giustizia.
Lo scopo del presente lavoro è una disamina delle potenzialità dell’Entomologia Forense e delle relative applicazioni come strumento spesso determinante per la risoluzione di intriganti e complessi casi giudiziari.
2. Introduzione
La classe degli Insetti(1) comprende circa un milione di specie ed è quindi la più numerosa tra quelle oggi studiate. Esistono numerosissimi ambienti di vita (habitats) nei quali è possibile rinvenire tali specie, essendo le loro capacità di adattamento e colonizzazione praticamente illimitate.
Un corpo in decomposizione è ovviamente compreso in tale insieme, costituendo una variegata ed appetibile sorgente di nutrimento nonché un luogo protetto ed adatto alla riproduzione. In letteratura sono riportate circa 400 specie colonizzatrici di corpi animali in decomposizione. Se da una parte è indiscusso il loro ruolo ecologico di degradatori della materia biologica, dall’altra essi rappresentano un utilissimo strumento per le indagini criminali come indicatori del luogo di rinvenimento di un corpo, per la ricostruzione di molti eventi e per la determinazione del P.M.I. (Post-Mortem Interval), il cosiddetto “intervallo post-mortem”.
Le tradizionali tecniche medico-legali attualmente utilizzate per il calcolo del tempo trascorso dalla morte di un individuo al ritrovamento del suo cadavere, si basano sul decremento della temperatura corporea, sullo studio del livor (macchie ipostatiche) e del rigor mortis (rigidità cadaverica) e sullo studio di elementi accessori come, ad esempio, il contenuto gastrico. Tali parametri, sebbene di fondamentale importanza nelle prime 24-48 ore, vanno irrimediabilmente incontro ad una stabilizzazione per la quale i dati ricavabili risultano poco o per nulla informativi (Greenberg and Kunich, 2002; Amendt et al., 2004) rispetto, invece, al dato entomologico che, secondo alcuni Autori (Kashyap & Pillay, 1989; Introna & Campobasso, 1998), almeno entro le 72 ore dalla morte rappresenterebbe il dato statisticamente più attendibile. Basti pensare, ad esempio, alla temperatura di un corpo umano ritrovato, in condizioni ambientali moderate, a 5-6 giorni dal suo decesso: tale parametro, da solo, non può fornire alcuna informazione precisa circa l’epoca della morte, potendo quest’ultima essere stimata indifferentemente tra il 2° (tempo necessario al naturale dissipamento del calore interno) ed il 6° giorno dal ritrovamento. Inoltre in alcuni casi il medico legale è impossibilitato a procedere a tali rilevazioni in sede di ritrovamento del cadavere, avendo a disposizione il corpo soltanto dopo la rimozione e il collocamento dello stesso presso le camere refrigerate degli obitori: in questo caso l’accurato utilizzo delle tradizionali tecniche perde significativamente la sua validità.
Per le ragioni sopra esposte lo studio della fauna entomologica e delle condizioni ambientali che ne hanno favorito lo sviluppo consentono oggi di ottenere delle stime tanatocronologiche di alta precisione (Introna e Campobasso, 1998), le quali, insieme alle risultanze analitiche prodotte da altre scienze forensi (es. antropologia, botanica, geologia, biologia, etc.), costituiscono il background tecnico scientifico determinante per la gestione ed il successo delle indagini.
Oltre al calcolo del P.M.I. il dato entomologico offre un importante spunto investigativo in relazione allo spostamento di un corpo e/o al suo occultamento, alla determinazione dei tempi di emersione dall’acqua, all’identificazione delle droghe e dei composti tossici che potrebbero aver causato la morte di un soggetto (la cosiddetta “Entomotossicologia”), fino ad arrivare ai casi di colpa professionale in ambiente ospedaliero ed omissione/negligenza nell’assistenza ai malati, applicando gli stessi criteri adottati per i reati di crimine violento.
a. Cenni storici
Ai giorni nostri la vista degli insetti genera paura o spesso disgusto ma, nella cultura delle civiltà più antiche, rappresentazioni di questi esseri sono state legate ad alcuni aspetti della vita e della morte fino ad essere persino considerati la personificazione di divinità.
La prima testimonianza dell’interesse dell’uomo verso il mondo degli insetti, e in particolare verso i Ditteri, si può trovare su un sigillo mesopotamico di 5000 anni fa in cui è rappresentata una gazzella distesa a terra apparentemente senza vita sopra la quale è raffigurata una mosca. L’attenzione verso l’insetto è probabilmente dovuta al fatto che questo periodo vide crescere in modo esponenziale l’insediamento umano, la quantità di rifiuti e le carneficine delle guerre e ciò ha avuto come conseguenza l’esplosione della popolazione di questi animali: “questi non poterono esser ignorati né dai vivi né dai morti”, essendo “i cadaveri uguali, siano essi di schiavi o di re” (Greenberg & Kunich, 2002).
Il primo chiaro riferimento alle mosche carnarie risale a più di 3600 anni fa, su una delle numerose tavolette di creta con scritti cuneiformi della “Har-ra-Habulla”, considerata a tutti gli effetti il più antico “libro di zoologia” conosciuto. Sono descritti 396 animali, tra i quali 10 mosche - incluse le mosche verdi (probabilmente Phaenicia sericata o Chrysomya albiceps) e le mosche blu (probabilmente il Genere Calliphora) -, entrambe, attualmente, di interesse medico-legale.
La relazione tra insetti e cadaveri era nota anche agli antichi Egizi: essi conoscevano il ciclo vitale dei Ditteri, compresa la metamorfosi, e incidevano amuleti a forma di mosca per scacciare il maligno e collane mortuarie che servivano ad allontanare la distruzione del corpo. Uno degli obiettivi dell’imbalsamazione del corpo era proprio proteggere il defunto dall’attacco degli insetti e del tempo: a tale proposito nel Capitolo 154 del Libro dei Morti si legge “Questo mio corpo non diverrà preda di larve”. Anche dopo l’imbalsamazione Ka, il guardiano dello spirito, rimaneva vicino al corpo o periodicamente tornava a controllarlo.
Nel papiro Gizeh n° 18026:4:14, trovato nella bocca di una mummia, si può leggere “Le larve non diventeranno mosche dentro di te”. Nelle epoche predinastiche compaiono amuleti di pietra a forma di mosca che avevano valore di prevenzione e protezione ma a partire dal Nuovo Regno la mosca assume anche significato di impudenza, persistenza e coraggio tanto da assegnare l’onorificenza dell’Ordine della Mosca Dorata o Mosca del Valore a soldati distintisi in battaglia.
Presso i Greci la mosca era considerata un animale sacro al quale si collegavano alcuni nomi di Zeus e Apollo: essa evocava l’onnipresenza degli Dei, incessantemente ronzanti, sempre in moto e pungenti. Ricordava anche il turbinio della vita olimpica e l’ideale di uomo d’azione, agile e febbrile, ma anche inutile e rivendicatore, così come il demone della Decomposizione, Eurinomo, divoratore di cadaveri, veniva rappresentato sia come avvoltoio sia come mosca. Il ruolo degli insetti nei fenomeni fisiologici della morte era ben conosciuta anche da Omero che immaginò Achille rivolgersi alla madre con tale preghiera: “Madre, son degne del divino fabbro, quest’armi, né può tanto arte terrena. Or le mi vesto, ma timor mi grava che nelle piaghe di Patroclo intanto vile insetto non entri, che, di vermi generator, la salma (ahi senza vita!) ne guasti sì, che tutta imputridisca”.“Pensier di questo non ti prenda, o figlio” - gli rispose la Dea - “l’infesto sciame, divoratore de’ guerrieri uccisi, io ne terrò lontano ov’anco ei giaccia intero un anno, farò sì, che il corpo incorrotto ne resti, e ancor più bello” (Omero, Iliade, libro 19, versi 20-33).
La relazione tra Ditteri e cadaveri si fece spazio anche in Asia minore dove, presso le civiltà dei Caldei, Filistei, Fenici ed Ebrei, le mosche erano associate a “Baalzebub”, Signore delle Mosche, rappresentato come una mosca che può portare piaghe di Insetti e mandare legioni alate come punizione (Greenberg 1991).
La prima testimonianza di una relazione “forense” mosca/omicidio viene fatta risalire ad un episodio accaduto in Cina, in un periodo compreso tra il 907 e il 970 d.C.: “un ufficiale della corte sentì una donna piangere e urlare disperatamente. L’ufficiale le chiese il motivo di questi lamenti e lei rispose che il marito era stato ucciso dal fuoco. L’ufficiale però si accorse che vicino al corpo si trovavano molte mosche e durante l’autopsia si scoprì una ferita sul capo dell’uomo. La donna confessò di aver ucciso il marito insieme a un altro uomo, procurandogli quella lesione” (Greenberg & Kunich, 2002).
Un altro “caso” risolto grazie ad evidenze entomologiche, seppur ancora ancestrali, viene fatto risalire al 1247 in Cina ed è riportato da Sung Tz’u nel suo “Hsi Yuan Chi Lu” (lo spazzare via i torti e lavare le ingiustizie), un primordiale, ma efficace, manuale per l’investigazione sulla scena del delitto: “Un lavoratore morì in un campo di riso, ucciso dai colpi di una falce. Il locale agente della legge si recò dove l’uomo era stato ucciso e radunò tutti i lavoratori dei campi di  quella zona. Chiese loro di deporre le loro falci in fila per terra. Il tempo era caldo e non ci volle molto tempo prima che le mosche iniziassero a raccogliersi su un falce particolare. Alla vista di tal evidenza il padrone della falce confessò il suo crimine”.
quella zona. Chiese loro di deporre le loro falci in fila per terra. Il tempo era caldo e non ci volle molto tempo prima che le mosche iniziassero a raccogliersi su un falce particolare. Alla vista di tal evidenza il padrone della falce confessò il suo crimine”.
Nelle opere d’arte del Rinascimento la morte e il demonio sono raffigurati come una mosca; si possono ricordare le opere del pittore veneziano del XV secolo Carlo Crivelli (“Madonna con Bambino”) e quella omonima di Giorgio Schiavone (1460), conservata nella Galleria Sabauda di Torino, dove si può vedere Gesù tenere in mano un uccellino, simbolo della Resurrezione, per proteggerlo da una mosca, simbolo di morte e di Satana (fig.1-2).
Un altro dipinto conservato a Torino, “San Gerolamo nel Deserto” di Matteo di Gualdo, ritrae il Santo in preghiera e dietro la sua figura si scorgono due volumi, chiusi con dei lucchetti, davanti ai quali è posata una mosca: il significato iconografico è che il Demonio, la mosca, non è in grado di toccare e disonorare le Sacre Scritture.
 Amboise Paré, uno dei più importanti medici militari del suo tempo, dà una descrizione personale della Battaglia di San Quintino del 1557: “vedemmo più di mezza lega di terra coperta da corpi morti; non potemmo resistere a lungo lì a causa dell’odore che emanavano i corpi morti e anche i cavalli morti. Pensai che noi ne eravamo la causa e vidi sciami di mosche intorno ai corpi che procreavano grazie all’umidità e si vedevano i loro corpi verdi e blu; erano così tanti da oscurare d’ombra il sole. Sentimmo il loro ronzio, che era ancora più orribile per noi. E pensai che fosse abbastanza per determinare una piaga, dove loro si posavano”.
Amboise Paré, uno dei più importanti medici militari del suo tempo, dà una descrizione personale della Battaglia di San Quintino del 1557: “vedemmo più di mezza lega di terra coperta da corpi morti; non potemmo resistere a lungo lì a causa dell’odore che emanavano i corpi morti e anche i cavalli morti. Pensai che noi ne eravamo la causa e vidi sciami di mosche intorno ai corpi che procreavano grazie all’umidità e si vedevano i loro corpi verdi e blu; erano così tanti da oscurare d’ombra il sole. Sentimmo il loro ronzio, che era ancora più orribile per noi. E pensai che fosse abbastanza per determinare una piaga, dove loro si posavano”.
Nel 1668 Francesco Redi compie il primo esperimento medico della storia seguendo il cosiddetto Metodo Scientifico Sperimentale: egli studia i fenomeni di decomposizione della carne sia esposta all’aria sia in recipienti chiusi. Con questo lavoro fu in grado di demolire sperimentalmente il concetto della “generazione spontanea” (abiagenesi), secondo cui le larve sarebbero prodotte “spontaneamente” dalla carne, teoria già enunciata da Aristotele e sostenuta fino ad allora dalla Scuola Aristotelica. A metà del diciannovesimo secolo un inventario delle specie di insetti e di altri artropodi legati ai fenomeni di decomposizione fu redatto da Orfila, ma senza un nesso con eventuali omicidi: in questo trattato, datato 1831, sono descritte 30 specie e viene preso in considerazione il ruolo dei Ditteri e dei Coleotteri nell’attacco e nella distruzione del cadavere, contribuendo anche ai primi studi sulla fauna riscontrata nelle esumazioni e sulla successione di artropodi che si avvicendano sul cadavere umano.
Solo nel marzo del 1850 si può ritrovare una prima documentazione di applicazione dell’Entomologia forense in occidente, quando Bergeret risolse un caso di infanticidio facendo uso degli insetti come indicatori del periodo di morte: il corpo di un bambino fu trovato murato nel camino di una casa e Bergeret determinò che l’insieme dei resti di insetti associati al cadavere (pupari di Musca carnaria) indicava uno stato di decadimento che risaliva a parecchi anni prima, per cui la responsabilità dell’infanticidio cadde sui precedenti occupanti della casa, non su quelli presenti al momento del ritrovamento (Bergeret, 1855). Negli anni successivi si verificarono casi simili a quello sopra citato e si ricorda l’intervento di Lazzaretti a Padova nel 1874: venne ritrovato un piccolo cadavere mummificato in una soffitta e per valutare l’epoca della morte vennero utilizzati i pupari e le larve che avevano colonizzato i resti.
È però nel 1894 che si data la nascita della moderna Entomologia forense, con la pubblicazione de “La faune des cadavres. Application de l’Entomologie a la Medicine Legale” di Jean-Pierre Mégnin. Egli determinò la sequenza e l’avanzamento della decomposizione di un corpo e osservò e descrisse i cambiamenti nei raggruppamenti di insetti col procedere del disfacimento del cadavere, definendo le “squadre di Artropodi” (Insetti e Acari) specifici di ogni stadio. Mégnin considerò il cadavere come un ecosistema dinamico con una sua particolare successione faunale, nonostante il corpo umano sia dimensionalmente finito. Mégnin pubblicò 14 lavori a partire dal 1883 e viene considerato dalla maggior parte degli autori come il padre dell’Entomologia forense, benché altri lo abbiano preceduto e molti dei suoi rigidi postulati siano ormai superati.
A partire del 1920 i medici legali italiani, con i contributi di Alessandrini, Bianchini e Porta cercano di razionalizzare i risultati degli anni precedenti, recuperando, non senza critiche, i concetti generali di Mégnin e apportando dati inediti sulla fauna cadaverica marina, grazie ad esperimenti orginali (Porta, 1929). Sempre in Italia, nel 1933, Bellussi pubblica un lavoro di accertamento della data di morte di quattro persone decedute nel deserto nordafricano, utilizzando l’entomofauna riscontrata durante il sopralluogo per effettuare esperimenti in condizioni controllate.
Gli studi del finlandese Nuorteva prima e quelli di Altamura e Introna poi, a cavallo tra gli anni ’70 ed ’80, hanno contribuito a chiarire il ruolo dei Ditteri cadaverici nella determinazione della cronologia della morte e hanno contribuito alla conoscenza morfologica degli stadi preimmaginali (cioè prima della fase adulta).
Nel 1986 Erzinçlioglu osserva che nell’Entomologia forense c’è una forma di conoscenza che non si può acquisire con la sola ricerca sperimentale, ma che richiede la raccolta delle analisi di casi pratici: questo giustifica la consuetudine da parte degli entomologi forensi di riportare, oltre agli aspetti scientifici della materia, anche la descrizione dei casi criminali affrontati e risolti mediante l’impiego di questa scienza. Negli stessi anni Marchenko descrisse i possibili casi di degenerazione dei tessuti dopo la morte: i processi della putrefazione sono dipendenti da moltissimi fattori quali età, sesso, costituzione, presenza/assenza di vestiti, tipo di copertura, luogo di morte, esposizione a fonti luminose, clima, substrato e molte altre condizioni. Tutte queste caratteristiche incidono sul corpo, ma anche su gli insetti che lo colonizzeranno, quindi decadono completamente i rigidi postulati di Mégnin che riguardano le successioni di ondate di insetti sul corpo nel tempo.
Tra il 1986 ed il 1989 K.G.V. Smith del British Museum di Londra pubblica i primi manuali di Entomologia Forense, ancora oggi considerati i testi di riferimento a livello mondiale. Infine durante l’XIX Congresso Mondiale di Entomologia (Pechino, 1990) venne per la prima volta organizzata una sessione autonoma di Entomologia Forense. In quella occasione i più importanti studiosi della materia (Nortueva, Goff, Lord, Haskell, Hall, Marchenko, Clark, Nishida e Wells) diedero vita ad un gruppo di lavoro composto da entomologi e medici legali, e furono così avviati incontri periodici e progetti di ricerca comuni, con scopi prevalentemente applicativi, tutt’oggi perseguiti da vari Enti europei.
3. L’Entomologia Forense
Nel XIII secolo vengono per la prima volta affiancati l’osservazione delle mosche con la presenza di un cadavere umano deceduto per cause non naturali: si viene così a definire l’iniziale profilo di un nuovo indirizzo di studio che mette in relazione la presenza dell’entomofauna cadaverica con casi di morte violenta o altri crimini, rendendo lo studio morfologico, ecologico ed etologico degli insetti applicativo nei casi medico-legali.
L’Entomologia Forense(2) è lo studio che comprende tutti gli aspetti applicativi degli insetti in qualche modo correlati a questioni di carattere legale: le conoscenze così ricavate possono essere utilizzate nelle indagini, nei processi penali e nelle controversie civili.
L’entomologia forense può essere suddivisa in tre branche principali (Lord & Stevenson, 1986; Catts & Goff, 1992):
a. Entomologia Urbana (Urban Entomology): procedimenti legali che coinvolgono insetti che influenzano manufatti dell’uomo e ambiente umano;
b. Entomologia dei Prodotti Immagazzinati (Stored Products Entomology): procedimenti giudiziari che coinvolgono insetti che infestano beni conservati e/o loro involucri (anche in casi di contrabbando) ed in particolare i prodotti alimentari (Entomologia Alimentare);
c. Entomologia Medico-Legale (Medicocriminal Entomology o Forensic Entomology): scienza che studia il coinvolgimento degli insetti in eventi di interesse tipicamente medico-legale (omicidi, suicidi, morti improvvise) e altri reati (abuso fisico, negligenze professionali).
Le informazioni che possono essere ricavate dalla raccolta e dalla determinazione degli Artropodi che si trovano sul luogo del reato sono molto importanti perché la presenza di questi animali non è mai occasionale: l’entomologia diventa quindi un nuovo e importante mezzo di indagine che può affiancare la secolare esperienza della medicina legale.
Inizialmente gli studi di Entomologia Forense vedevano in modo complessivo il fenomeno della decomposizione, con grande attenzione verso l’aspetto ecologico; col tempo si è cercato di approfondire la conoscenza delle specie che fanno capo a questo processo, senza dimenticare il loro ruolo nell’ecosistema ma cercando di raccogliere i dati in modo utile alle applicazioni forensi.
Negli ultimi decenni l’evoluzione della materia ha voluto seguire con maggior impegno la strada dello studio approfondito della biologia delle specie di maggiore interesse forense (morfologia, chiavi dicotomiche, sviluppo, cicli di vita) attraverso l’utilizzo delle tecniche analitiche più moderne e la ricerca di un metodo di lavoro ottimale e risolutivo.
Il quadro in cui possono essere inserite le attuali linee di ricerca applicata comprende:
- individuazione delle specie coinvolte nel decadimento dei cadaveri;
- perfezionamento delle chiavi dicotomiche (cioè degli strumenti di classificazione delle specie) e definizione di nuove;
- studio dell’ecologia e dell’etologia delle specie interessate;
- estensione sistematica dello studio ad ambienti diversi;
- perfezionamento e ricerca di criteri affidabili per il calcolo dei tempi di sviluppo, per la determinazione degli stadi di vita degli insetti e dell’influenza della temperatura sulla velocità di sviluppo;
- applicazione di metodologie chimiche e biologico-molecolari al tradizionale esame morfo-fisiologico degli insetti;
- definizione di criteri per la gestione dei casi pratici, con la messa a punto di “checklist” e protocolli per il sopralluogo, il repertamento ed il trattamento dei campioni.
Partendo dalle ricerche pubblicate in letteratura, alcuni autori hanno raccolto dati riguardanti solo alcune delle specie di interesse medico legale (soprattutto a quelle dei generi Calliphora, Protophormia terranovae, Lucilia e Cochliomyia) ma la gran parte del lavoro deve ancora essere sviluppata a causa della esigua quantità di specie repertate, della scarsezza di dati scientifici necessari per disporre di una raccolta sistematica e del mancato approccio multidisciplinare alle linee di studio sopra elencate.
a. L’entomofauna necrobionte
Da un punto di vista strettamente scientifico, la carcassa animale o il cadavere umano può essere considerato un vero e proprio piccolo ecosistema, che compare improvvisamente nell’ambiente e che rappresenta un’importante risorsa per numerosi organismi: mammiferi, pesci, uccelli, invertebrati, vegetali, funghi e batteri si trovano a disporre di una inaspettata fonte alimentare che non pone resistenza ad essere predata e che viene disputata tra i consumatori/degradatori che competono tra loro per esigenze alimentari affini.
Come per tutti gli ecosistemi di nuova formazione, la fauna(3) presente su una carcassa si costituisce dapprima tramite la colonizzazione ad opera di organismi detti “pionieri”, che cominciano a modificare l’ambiente, mentre in seguito viene incrementata da altri organismi con un ordine temporale ben preciso dipendente delle loro esigenze di nicchia ecologica.
Come l’ambiente progressivamente si modifica, le specie cambiano oppure permangono in uno stato di equilibrio sia con il substrato che li circonda sia con le altre specie competitrici, predatrici, commensali o parassite.
A differenza di altri ecosistemi costituiti da materiale non vivente, quello cadaverico è un ecosistema effimero, che appare improvvisamente e che scompare altrettanto rapidamente, consumato dai suoi stessi decompositori fino a che non resta più nulla di “commestibile”, eccezion fatta per quei tessuti che non vengono affatto degradati (ossa).
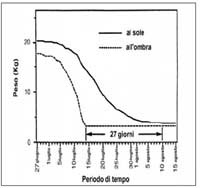
La fauna saprofaga(4), analizzata in termini quali-quantitativi e correlata all’ambiente di ritrovamento, al clima, al tempo e alle fasi di decomposizione cadaverica, è un utile indizio che permette di risalire al momento in cui è comparso il corpo in una determinata zona, quindi al momento del decesso ma anche all’eventuale percorso fatto dal cadavere prima del rinvenimento. In alcuni casi è possibile pervenire a conclusioni che riguardano la modalità della morte e l’identificazione personale. Le possibili ricostruzioni per “via zoologica” degli eventi riconducibili ad un cadavere si basano soprattutto sullo studio della micro-fauna cadaverica facilmente rinvenibile sul corpo. Per ciò che concerne la macro-fauna (ratti, topi, piccoli predatori, uccelli, pesci,) essa può fornire informazioni per lo più accessorie (es.: lesioni di morsi di diverse specie sono utili per descrivere i percorsi seguiti dal cadavere dopo la morte). L’insieme dei cambiamenti introdotti su una carcassa dalla presenza di piccoli invertebrati può essere, infatti, molto più significativa dei cambiamenti arrecati da animali di taglia superiore. Payne (1965) dimostrò l’incisività degli artropodi nei processi di consunzione cadaverica registrando perdite di circa il novanta per cento del peso iniziale della cavia dopo solo una settimana d’esposizione alla colonizzazione dei Ditteri, mentre registrò una perdita dell’ottanta per cento della massa, dopo 6/7 settimane, in cavie non esposte all’azione della microfauna.
Della gran quantità di organismi che fanno parte della fauna necrobionte(5), il gruppo più rappresentato a livello qualitativo (varietà di specie) e quantitativo (presenza di esemplari) è quello degli Artropodi Esapodi (Insetti).
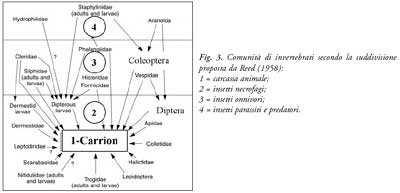
Per quanto riguarda gli insetti necrobionti, si possono definire quattro categorie ecologiche rinvenibili all’interno della comunità presente (Smith, 1986; Catts & Goff, 1992) sia contemporaneamente che in tempi diversi sul substrato in decomposizione (fig. 3):
a. necrofagi spazzini: soprattutto Ditteri, si nutrono delle carcasse di organismi animali;
b. parassiti o predatori dei necrofagi: Ditteri, Coleotteri, acari, ragni e altri artropodi. In alcuni casi gli insetti sono necrofagi durante i primi stadi di sviluppo e diventano predatori negli ultimi stadi;
c. onnivori: vespe, formiche e alcuni coleotteri, specie opportuniste che si nutrono sia del corpo che dei suoi colonizzatori;
d. occasionali o criptozoici: acari, ragni, formiche, collemboli e chilopodi. Vivono nell’ambiente circostante e si nutrono normalmente d’altro, ma possono trovarsi occasionalmente sul cadavere, ivi trasportati dal vento o attratti dal calore e dall’umidità del corpo.
L’insetto necrofago(6) utilizza i propri organi di senso altamente specializzati per individuare il substrato cadaverico sin dalle fasi iniziali post-mortali, quando la degradazione della materia determina lo sviluppo di gas e odori particolari, generati dall’esaurimento della materia organica. Grazie alle proprie caratteristiche fisiche e fisiologiche gli insetti sono i primi organismi che trovano e iniziano a modificare un substrato organico morto, sia esso carogna animale o derrata vegetale: allo stesso modo la degenerazione del corpo umano in condizioni naturali è accompagnata dalla presenza d’insetti necrofagi.
Gli insetti attratti da resti in via di decomposizione sono un numero molto ampio, oltre 400 specie (Payne, 1965), ma i gruppi che giocano un ruolo attivo nel processo di decadimento sono in particolare due: i Ditteri (Diptera) e i Coleotteri (Coleoptera), in altre parole i necrofagi ed i loro predatori (Leclercq & Verstraeten, 1993).
In questo lavoro è stato scelto di focalizzare l’attenzione sull’entomofauna necrobionte, ma non bisogna comunque dimenticare che sul substrato in disfacimento è presente anche una massiccia fauna non specializzata nella decomposizione delle carcasse (Anderson & Van Laerhoven, 1996), che è stata spesso trascurata dagli specialisti, nonostante queste specie siano particolarmente importanti come partecipanti occasionali attivi alla degradazione del corpo. Essi possono anche agire come potenziali competitori, predatori e parassiti della fauna cadaverica, in grado di inibire in parte o del tutto lo sviluppo di alcune specie normalmente utilizzate come principali indicatori dei tempi di morte.
Piccoli predatori (ratti, topi, corvi, etc.) possono nutrirsi del cadavere, senza spostare in modo apprezzabile i resti ma lasciando particolari segni sul corpo. I roditori possono rosicchiare sia la carne che le ossa apportando lesioni nettamente distinguibili.
In alcune condizioni i grandi predatori (alligatori, orsi, avvoltoi, cani, etc.) possono influire sulla decomposizione del cadavere, cibandosi del corpo e disperdendone le parti.
Se il corpo viene rinvenuto in stadio di decomposizione avanzato ma illeso, in un’area in cui è probabile la presenza di vertebrati di piccola e media taglia, è verosimile ritenere che esso sia rimasto in un luogo dove i possibili predatori non avevano accesso.
Altri insetti come formiche (ma anche blatte e vespe) possono cibarsi delle parti esterne del corpo, lasciando effetti superficiali molto caratteristici, piccoli crateri poco profondi, di forma e in posizioni irregolari.
Durante il Convegno EAFE (European Association for Forensic Entomology) del 2004, svoltosi in Londra, fu presentata un’interessante relazione, basata su casi reali, che focalizzava l’attenzione sull’effetto dell’attività delle formiche sulla velocità di decomposizione dei corpi e sull’interpretazione della lesività prodotta da questi Imenotteri.
Alcune abrasioni cutanee causate da formiche possono, infatti, essere facilmente confuse con lesioni prodotte in vita. In realtà, si tratta di modificazioni superficiali dell’epitelio prodotte sia dall’azione meccanica delle formiche intente a cibarsi dei frammenti cutanei, sia dall’azione caustica dell’acido formico secreto dagli Imenotteri stessi.
Tra i casi illustrati degno di nota è quello del cadavere di un minore diffusamente interessato da lesioni post-mortali da formiche, in particolare al collo, ove, nonostante l’azione degli imenotteri, fu ancora possibile rilevare alla ispezione cadaverica gli esiti di manovre d’afferramento e soffocamento, causa del decesso (Campobasso et al., 2004).
Grandi e piccoli predatori, possono inoltre, direttamente o indirettamente, disturbare l’entomofauna nutrendosi di essa e lasciando sul cadavere una biocenosi(7) del tutto diversa da quella che ci sarebbe in condizioni normali: questa eventualità può modificare in modo importante il decorso della decomposizione e rendere molto complessa la stima dell’intervallo post-mortem sia in ambiente terrestre sia in ambiente acquatico (Magni P. & Scaravelli D., 2006).
b. Ditteri e coleotteri di interesse forense
Gli insetti attratti da resti in via di decomposizione sono un numero molto ampio, ma i gruppi che giocano un ruolo attivo nel processo di decadimento sono in particolare due: i Ditteri e i Coleotteri.
1. I ditteri
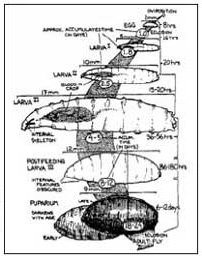
I ditteri (Ordine Diptera)(8) sono tra gli insetti più numerosi e presenti nella biosfera terrestre. Il ciclo di sviluppo di questi animali prevede una metamorfosi completa uovo-larva-adulto con grandi differenze strutturali ed ecologiche tra gli stadi giovanili e l’immagine (adulto), nella quale si trasformano dopo un periodo trascorso all’interno di una “pupa”.
Le uova vengono generalmente depositate sul cadavere in siti con facile accesso alle cavità interne e con un certo grado di umidità (orifizi naturali del corpo), sono molto piccole e spesso concentrate in piccoli cumuli.
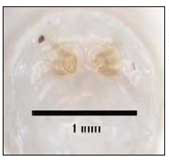
Le larve (fig. 4-5), che appaiono come piccoli vermetti bianchi, presentano almeno tre stadi di sviluppo la cui distinzione è possibile grazie a particolari caratteristiche morfologiche osservabili con l’ausilio di un microscopio ottico.
Inglobata una quantità sufficiente di cibo, le larve smettono di mangiare (larve in post-feeding) e generalmente si allontanano in cerca di un luogo sicuro dove “impuparsi”, cioè dove costituire l’involucro (pupa) all’interno del quale avverrà la metamorfosi in insetto adulto.
Lo spostamento delle larve in post-feeding può variare da zero a 50 metri, alcune specie tendono a impuparsi direttamente sul pabulum e altre a scavare nel terreno (Fuller, 1934; Norris, 1959; Turner, 1991): la conoscenza di queste abitudini diventa di estrema importanza durante il sopralluogo entomologico.
La pupa (fig. 6) rappresenta lo stadio transitorio necessario per la metamorfosi nello stadio adulto.
Lo farfallamento dell’immagine ha una durata variabile compresa da pochi minuti a un’ora, sempre in dipendenza dalle condizioni ambientali; il regime alimentare di questi insetti si basa su una varietà molto grande di sostanze, dai prodotti della decomposizione al nettare fino al sangue, usato prevalentemente dalle femmine per il suo contenuto di proteine necessarie alla produzione delle uova.


I Ditteri vivono in stretto contatto con l’uomo e costituiscono veicoli i trasmissione per infezioni e malattie anche degli animali domestici. In particolare alcuni inducono debilitazione nell’uomo e negli animali domestici con infestazioni che prendono il nome di “miasi”. La miasi è un’infestazione cagionata da larve di Ditteri a parassitismo obbligato o accidentale che per un tempo variabile si alimentano su tessuti vivi o necrotici dell’ospite, dei liquidi o degli alimenti ingeriti (Zumpt, 1965). Gli insetti “miasigeni” sono di grande importanza in Entomologia forense perché la loro presenza su un corpo umano in vita è spesso sinonimo di incuranza nell’igiene: sono spesso gli anziani e i bambini (Pikles et al., 2005) a presentare questo tipo di parassitosi perché sono soggetti che dipendono da tutori la cui affidabilità professionale può essere oggetto di indagine da parte del medico legale e dell’entomologo forense.
Da un punto di vista più criminalistico è interessante notare che, secondo alcuni medici legali, le lesività prodotte dalla massiccia fuoriuscita epidermica delle larve in miasi potrebbero, se osservate su tessuti in avanzato stato di decomposizione (quindi in assenza delle caratteristiche ulcerazioni miasiche), assomigliare alle ferite prodotte da un arma da fuoco con munizionamento “ a pallini” (le cosiddette “rosate”). L’attenta analisi permetterebbe però di desumere la direzionalità dell’azione lesiva dai margini delle ferite, i quali saranno ovviamente rivolti verso l’esterno nel caso di pregressa miasi (Catts and Haskell, 1990).
L’attacco del cadavere da parte dei Ditteri avviene piuttosto precocemente, appena l’iniziale odore della putrefazione diviene percettibile al sensibilissimo apparato olfattivo di questi insetti: la colonizzazione diventa pressoché immediata durante la stagione estiva, quando le emanazioni sono più forti ed è presente un maggior numero di Ditteri attivi.
Sebbene fortemente attratti dai prodotti della putrefazione, i Ditteri sono capaci di colonizzare un cadavere solo se è accessibile e non sotterrato: bastano 2,5 cm di terra per impedire l’attacco delle specie dominanti tra i Ditteri della prima ondata, con il conseguente rallentamento della decomposizione dei resti interrati (Lundt, 1964). Alcuni Ditteri dei generi Muscina (Famiglia Muscidae), Metopina e Conicera (Famiglia Phoridae) sono in grado di colonizzare corpi sepolti, deponendo uova sul terreno in risposta alla presenza di liquami organici o all’emanazione di gas putrefattivi: le larve che si nutrono dei materiali presenti in superficie dopo aver raggiunto il secondo stadio possono penetrare lentamente in un terreno morbido e raggiungere la carcassa (Mégnin, 1894; Lundt, 1964; Payne, 1965; VanLoerhoven & Anderson, 1999).
I Ditteri depongono generalmente in parti del corpo come narici, occhi, meati dell’orecchio, cavo orale, vagina e retto, zone che mantengono un certo grado di umidità e che facilitano l’accesso verso l’interno, ma l’eventuale presenza di ferite o lacerazioni può garantire una scelta più ampia.
La dimensione del cadavere può limitare la disponibilità di cibo, quindi influenzare la sopravvivenza delle larve che si alimentano su e di esso e il numero di cicli che possono susseguirsi sul substrato. Inoltre, ulteriori fattori che assumono valore fondamentale per il successo del completamento del ciclo vitale sono:
- la competizione intra ed interspecifica (cioè rispettivamente tra individui della stessa o di specie diverse)(Smith, 1986);
- i fenomeni di diapausa (interruzione dello sviluppo con durata temporale prestabilita) e quiescenza (interruzione dello sviluppo con durata dipendente dal ripristino delle condizioni ambientali favorevoli);
- le eventuali predazioni ed i parassitismi delle larve e delle pupe dei Ditteri da parte di altri insetti (es. Imenotteri).
La conoscenza del comportamento delle specie è indispensabile per l’analisi interpretativa della presenza dei Ditteri e degli altri insetti sulla scena del crimine. L’attività, la competizione e la possibilità di questi insetti di bloccare il corso della loro crescita sono fattori determinanti per lo studio della biocenosi cadaverica al fine di stimare il PMI e altri parametri.
2. I coleotteri
I coleotteri (Ordine Coleoptera)(9) rappresentano l’ordine con il maggior numero di specie, costituendo il 40% di tutti gli insetti conosciuti. Sono caratterizzati dall’avere il paio d’ali anteriori sclerotizzate, dette “elitre”, e il paio d’ali posteriori, se presenti, membranacee e ripiegate sotto le prime. Sono insetti che subiscono una metamorfosi completa, le forme larvali sono molto varie e le specie sono sovente di difficile determinazione. All’interno di quest’Ordine si trovano molti gruppi d’interesse forense, ma a causa dei diversi ruoli giocati dalle varie specie di Coleotteri non esiste un momento caratteristico in cui gli individui compaiono durante il processo di decomposizione. Generalmente tendono a colonizzare i cadaveri più tardi dei Ditteri e la loro presenza è in un numero più limitato.
I Coleotteri possono nutrirsi direttamente del cadavere (Silfidi, Dermestidi, Nitidulidi e alcuni Cleridi) oppure essere predatori di Ditteri. A volte si trovano casualmente anche specie fitofaghe o polifaghe (specie che si nutrono di materiale vegetale o d’altra natura), ma queste non sono da considerarsi importanti ai fini medico-legali a meno che la loro presenza non indichi uno spostamento del corpo da un ambiente all’altro.
Nonostante i tempi di sviluppo di gran parte dei Coleotteri siano meno conosciuti rispetto a quelli dei Ditteri, il confronto con questi può servire a confermare o perfezionare la stima del P.M.I., definendo la finestra temporale al termine della quale le mosche hanno completato il ciclo vitale ed abbandonato il cadavere (Bellussi, 1933).
c. Il ruolo dell’entomologo forense
Il ruolo dell’Entomologo forense nell’ambito delle indagini su crimini violenti è quello di riuscire a correlare la presenza della biocenosi campionata sul substrato cadaverico con i parametri ambientali specifici del luogo di morte per poter stimare con sufficiente precisione:
- la datazione dell’epoca della morte o P.M.I.;
- il luogo del decesso ed eventuali spostamenti;
- la correlazione degli insetti con la verosimile fonte di nutrimento;
- gli eventi succedutisi prima del ritrovamento del corpo;
- le eventuali lesioni che potrebbero essere la causa di morte;
- l’eventuale consumo di sostanze stupefacenti;
- l’eventuale esposizione a sostanze tossiche.
In Italia la figura professionale dell’Entomologo forense è pressoché sconosciuta e poco impiegata, mentre da parecchi decenni è diventata irrinunciabile in molti Paesi come gli Stati Uniti. Il Federal Bureau of Investigation (F.B.I.), prima Agenzia governativa ad investire massivamente sulle potenzialità delle scienze forensi, istituì alcuni anni fa una struttura specializzata nello studio degli insetti presso il “Forensic Science Research and Training Center” di Quantico (Virginia) e sono in crescita i corsi di formazione del precipuo settore anche grazie alla diffusione mediatica. Nella nota serie televisiva “C.S.I.”, infatti, il detective Grissom ricopre il ruolo di Entomologo Forense a capo di una squadra di specialisti continuamente impegnati nella risoluzione di intrigati casi giudiziari. Il recente bestseller della nota scrittrice e criminalista Patricia Cornwell, “Calliphora”, ha come personaggi principali dei ditteri (Calliphoridae) rinvenuti sul luogo del delitto, così come il recente successo editoriale della stessa autrice “La Fabbrica dei Corpi” è invece ispirato ad un centro di ricerca realmente esistente nel Tennessee (U.S.A.), conosciuto appunto come “The Body Farm”, in cui vengono testate tutte le condizioni possibili inerenti alla decomposizione fisiologica ed entomologica dei cadaveri umani.
In Europa la Gendarmeria francese ha avviato, già da qualche anno, il “Départément Entòmologie” presso l’“Institut de Réchèrche Criminelle de la Gendarmèrié” mentre l’analogo laboratorio della Polizia Scientifica Svizzera è attivamente indirizzato nello studio e nell’applicazione di protocolli di sopralluogo ed analisi di campioni entomologici. Il Forensic Science Service inglese, uno tra i più autorevoli enti a livello mondiale nel campo delle investigazioni scientifiche, ha già da tempo formato i propri esperti in entomologia forense. Altri paesi europei presentano complessivamente una situazione ibrida in cui tali competenze sono delegate sia a strutture universitarie (Dipartimenti di Entomologia Forense presso Istituti di Medicina Legale) sia a laboratori specializzati delle forze dell’ordine. Recentemente anche la Gendarmeria Reale del Marocco ha voluto dotarsi del “Départment Entòmologie Judiciaire” all’interno del “Laboratòire de Réchérche et d’Analyses Techniques et Scientifiques”.
Negli ultimi anni si è verificato un sostanziale rinnovamento nella strumentazione e nelle tecniche utilizzate nelle applicazioni medico-legali, lo sviluppo delle quali può così elargire un contributo più ampio della sola datazione del P.M.I., peraltro sempre più precisa grazie all’utilizzo di sistemi informatico-statistici (Schoenly et al. 1996; Von Zuben et al., 1996). L’attività dell’entomologo forense in Europa ha iniziato ad essere una realtà importante anche grazie ai progressi che nel tempo si sono avuti con l’ausilio di tecniche moderne ed efficaci come le indagini biochimiche e molecolari a fianco della metodica classica di studio. D’altra parte molto deve essere ancora fatto nel nostro Paese affinché tutti gli operatori del settore forense siano perlomeno portati a conoscenza delle potenzialità di tale approccio.
d. La datazione dell’ epoca di morte: calcolo del P.M.I.
La datazione dell’epoca della morte (Post Mortem Interval, P.M.I.) è il parametro principale che l’Entomologo forense deve essere in grado di identificare. Per poterlo stimare si utilizzano fondamentalmente due approcci (Haskell et al., 1997; Erzinçlioglu, 1983):
a. lo studio della natura della biocenosi presente sul cadavere, metodo utilizzato per studiare un P.M.I. lungo e dove intervengono più ondate successive di insetti necrofagi;
b. la determinazione dello stadio di vita degli insetti che vengono rinvenuti sulla carcassa, metodo utilizzato per studiare un P.M.I. breve e dove i Ditteri della prima ondata sono protagonisti.
In entrambi i casi il luogo di ritrovamento del corpo e le temperature ambientali sono le informazioni di base per la corretta interpretazione dei dati entomologici. Tuttavia, l’analisi di questi ultimi non prescinde mai dall’esame medico legale sulla decomposizione cadaverica ma è di supporto a questa: se la fase di decomposizione del corpo non corrisponde alle specie ed alle fasi vitali degli insetti rinvenuti, l’investigatore dovrebbe sospettare che il processo è stato interrotto o modificato in qualche modo (Catts & Haskell, 1990).
L’analisi della natura della biocenosi comprende l’identificazione delle specie alla luce dei fenomeni trasformativi post-mortem e delle caratteristiche ambientali.
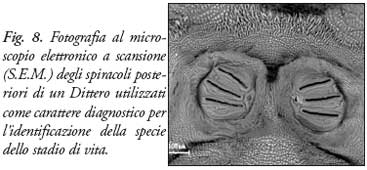
Identificazione di Specie - Per definire univocamente una specie si ricorre a metodi basati sulle chiavi dicotomiche, cioè al confronto con connotazioni morfologiche, riportate ed illustrate nei compendi entomologici, che permettono l’attribuzione di un dato esemplare ad una specie ben caratterizzata. L’osservazione di tali strutture può essere eseguita con il microscopio ottico, lo stereoscopio ma anche con il microscopio elettronico a scansione (S.E.M.). è oggi possibile utilizzare anche l’analisi del DNA per stabilire l’appartenenza di un insetto ad una data specie. Una volta caratterizzata la specie si considerano i fenomeni trasformativi e l’ambiente circostante. I fenomeni trasformativi post-mortem nei diversi ambienti costituiscono invece una delle variabili più influenti nel calcolo del P.M.I.

Fenomeni Trasformativi - La putrefazione rappresenta il più importante processo naturale di distruzione post-mortale della materia organica. L’entomologo forense deve essere pienamente edotto circa l’insieme dei cambiamenti fisici e biochimici cui il corpo va incontro dopo la morte. Questi sono didatticamente suddivisi, dalla medicina legale, nelle seguenti fasi o “stadi di decomposizione”:
- stadio fresco (fresh stage);
- stadio cromatico (cromatic stage);
- stadio enfisematoso o gassoso (bloated stage);
- stadio colliquativo (decay stage e advanced decay);
- stadio della scheletrizzazione (dry stage).
I fenomeni trasformativi determinano profonde modificazioni dell’aspetto e della struttura del cadavere che sono caratteristiche per ognuna delle fasi appena elencate; essi non devono essere intesi come momenti rigidamente individuabili, quanto come una sequenza di fenomeni che si sovrappongono e si combinano tra loro in una progressione ininterrotta di eventi, fino alla distruzione della materia organica (Introna & Campobasso, 1998).
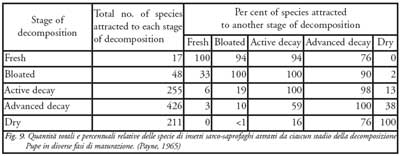
Ovviamente ogni stadio presenta una sua caratteristica popolazione entomologica (fig. 9) la quale, però, può essere non esclusiva di una data fase e comunemente associata alla presenza di altre specie. Anche in questo caso le condizioni ambientali giocano un ruolo fondamentale influenzando contemporaneamente sia il naturale fenomeno putrefattivo, causato dall’azione enzimatica e dei microrganismi, sia l’azione della micro-fauna necrobionte. D’altra parte le specie d’insetti presenti sui resti organici dipendono fortemente dal particolare ambiente in cui è deposto il corpo e dalle modificazioni dello stesso. Quindi se da una parte l’ambiente seleziona la fauna cadaverica, è altresì vero che quest’ultima modifica l’ambiente stesso di crescita.
Caratteristiche Ambientali. Alla luce di quanto esposto si può affermare che l’enorme quantità di habitat esistenti e delle eventuali modificazioni rende praticamente impossibile una conoscenza analitica e sistematica di tutte le situazioni reali che si possono presentare; uno studio sugli ambienti più tipici può invece fornire degli importanti riferimenti di base.
a. Corpi in ambiente terrestre.
L’ambiente terrestre può causare profonde deviazioni dai valori riportati nelle “tabelle” in considerazione dei molti fattori che incidono sui fenomeni tanatologici. Tra i più importanti ricordiamo: clima e microclima (fig. 10), altitudine e latitudine, presenza e tipo di vegetazione, presenza di animali in grado di modificare l’habitat, presenza di acqua (dolce, salata o paludosa), natura del terreno (natura chimica, permeabilità, compattezza, etc.), ambiente rurale o urbano, ambiente coltivato (i pesticidi determinano spesso un abbattimento differenziale dell’entomofauna, per cui molte specie che si trovano negli ambienti circostanti non si trovano più nelle coltivazioni;
b. Corpi in ambiente chiuso.
I luoghi più comuni sono le abitazioni con porte e finestre serrate, i bagagliai degli automezzi, i sacchi di plastica, etc.: più l’ambiente è “ermetico” più sarà difficile per gli insetti raggiungere il corpo e deporre le uova. Bisogna tenere presente che sono animali piuttosto piccoli e basta una piccola fessura per permettere loro l’entrata. D’altra parte la ricerca di una via d’accesso al cadavere può portare ritardi che non sempre è possibile quantificare e che riducono il calcolo del P.M.I. alla stima di un P.M.I. minimo. Inoltre gli ambienti chiusi e/o quelli sottoposti a massiccia disinfestazione, al pari degli ambienti sottoposti all’uso intensivo di pesticidi, attuano una selezione dell’entomofauna. L’occultamento di un corpo in spazi inaccessibili o con sistemi di copertura a tenuta ermetica “cristallizzerà” l’entomofauna eventualmente presente al momento dell’occultamento del cadavere, avendo l’isolamento fisico impedito l’accesso di altre specie delle “ondate successive”;
c. Corpi Sepolti.
Il seppellimento, in dipendenza della profondità, rallenta il processo di decomposizione e ostacola o impedisce l’accesso agli invertebrati, modificando la normale successione delle specie. Solo un’entomofauna limitata raggiunge il corpo: questi insetti depongono le uova sulla superficie del terreno e le larve, attratte dal cadavere, lo raggiungono scavando nel terreno. Il processo di decadimento di un corpo sepolto, quindi, è simile a quello di un corpo esposto, cambia solo il tipo di fauna; la presenza di insetti caratteristici della superficie, quindi, deve far supporre che la sepoltura sia avvenuta in un tempo successivo alla morte e alla colonizzazione da parte di Esapodi tipici dell’habitat esterno. In particolare solo alcuni Calliforidi depongono uova sul terreno, i Muscidi comprendono larve che migrano fino a 2,5-10 cm di profondità ed infine gli adulti d’alcuni Foridi (Conicera e Metopina) e i Coleotteri Stafilinidi del genere Athena, grazie alle loro dimensioni ridotte, possono raggiungere e colonizzare i cadaveri interrati anche a maggiore profondità (Lundt, 1964). La possibilità d’accesso al corpo dipende anche dal tipo di terreno, ad esempio in un terreno compatto e impermeabile come quello argilloso gli insetti possono accedere alla materia in decomposizione solo qualora la copertura di terra sia rimossa dagli animali o lavorata dall’uomo (Turner & Wiltshire, 1999). Più alto è lo strato di terreno sovrastante il cadavere, più lento sarà il processo di decomposizione e minore sarà il numero di insetti necrofagi che lo raggiungono (Turner, 1991). Studi condotti su cadaveri da Rodriguez e Bass (1985) hanno permesso di verificare che dopo un anno di sepoltura ad una profondità di un metro è possibile trovare grandi quantità di tessuto ma non si trovano insetti. Il problema dei cadaveri sepolti è molto complesso e non di facile soluzione: esiste solo un unico studio sperimentale sulla biocenosi dei corpi interrati nel quale vengono considerate solo la velocità di decadimento dei cadaveri e le caratteristiche del suolo (Motter, 1898). Ancora oggi non esistono modelli esaurienti sulla relazione tra tipo di suolo e azione degli insetti sul corpo inumato;
d. Corpi in ambiente acquatico.
L’ambiente delle acque dolci e salate, in cui si trovano numerose larve di insetti, presenta una “nuova frontiera” per l’Entomologia forense (Haskell et al., 1997; Magni e Scaravelli, 2006;) dal momento che non esistono insetti puramente necrofagi confrontabili con le specie terrestri. Il corpo immerso in acqua dolce o salata subisce un processo di decomposizione diverso rispetto a quello tipico dei cadaveri esposti all’aria, questo a causa dell’azione del liquido sul corpo e della diversa azione della fauna cadaverica che lo attacca. Gli insetti che abitano l’ecosistema acquatico non hanno abitudini alimentari esclusivamente necrofaghe come capita per le specie terrestri, gli esapodi e i vertebrati acquatici vengono in contatto con il cadavere in qualità di opportunisti, predatori diretti/indiretti o semplicemente in modo casuale. Quando il corpo è completamente sommerso, la deposizione di uova da parte degli insetti della fauna terrestre è impossibile. Quindi il ritrovamento di uova di artropodi terrestri deve suggerire l’occultamento post-mortem per immersione. Cambiando prospettiva è anche possibile ipotizzare che insetti caratteristici degli ambienti acquatici (fiumi, stagni e paludi) siano rinvenuti su un cadavere rinvenuto molto distante da tali habitat (fig. 11); anche in questo caso si può immaginare che il corpo sia stato spostato dal luogo del evento criminoso.
Anche l’utilizzo di alghe, depositi di sedimenti e la presenza di macroinvertebrati acquatici sia come animale integro sia come gusci, pupari e/o frammenti di entrambi hanno dimostrato la loro utilità come mezzi di investigazione. Sebbene nel contesto acquatico esistano pochi indicatori entomologici con una precisione comparabile a quella degli insetti terrestri, sono comunque possibili delle osservazioni che aiutano nella stima del tempo d’immersione dopo la morte (PMSI, Post-Mortem Submersion Interval), come ad esempio le fasi di sviluppo d’animali e piante marine che usano il corpo come substrato di crescita.
I metodi utilizzati nell’analisi dello stadio di vita degli insetti generalmente comprendono un confronto tra i campioni prelevati sulla scena del crimine con campioni noti allevati in condizioni controllate (Kamal, 1958; Nortueva, 1977; Smith, 1986; Williams, 1984).
È possibile stimare l’età del campione in esame riconoscendo alcune caratteristiche morfologiche e fisiologiche che distinguono gli stadi dello sviluppo dall’uovo all’adulto (fig. 12). In seguito si procede, per ottenere una stima più precisa, al sezionamento ed all’esame microscopico dei tessuti dell’esemplare (in particolare la larva ed in alcuni casi della pupa).
Alcuni parametri come la sovrapposizione di strutture dello stadio “nascente” visibili sotto quelle dello stadio precedente (fig. 13) o il cambiamento di colore (fig. 14) delle pupe vengono attentamente considerati per stabilire lo stadi larvale e l’età del campione.
Esistono diverse procedure attraverso cui è possibile definire l’età di una larva (Amendt et al., 2004):
- calcolo della lunghezza o del peso secco della larva e la comparazione di queste stime con dati di riferimento;
- studio delle strutture microscopiche;
- stima della storia termica dei Ditteri, o somma termale.


La lunghezza e il peso della larva sono dei parametri direttamente proporzionali che crescono durante il periodo di feeding (nutrimento) e decrescono nel periodo post-feeding (Nishida, 1984; Davies & Ratcliffe, 1994); l’utilizzo di un “isomegalediagramma” e delle scale di maturazione proposte in letteratura risultano fondamentale per la corretta identificazione dello stadio dei campioni rinvenuti, poiché l’accorciamento delle larve, nel periodo post-feeding, può sfalsare la stima. Il calcolo del periodo di tempo necessario allo sviluppo dei diversi stadi larvali è possibile grazie a studi in laboratorio condotti su diverse specie di insetti necrofagi, in condizioni controllate (temperatura, luce e umidità) e direttamente sul campo in condizioni naturali selezionate (corpo esposto alla luce, in ombra, sotterrato, in acqua, bruciato, etc.). Queste sperimentazioni hanno permesso di stilare dei grafici che rappresentano un ottimo gradino di partenza per l’analisi dei nuovi casi, ma che non devono limitare l’Entomologo forense ad una mera opera di applicazione dei valori espressi in essi. Ogni caso, infatti, è caratterizzato da variabili peculiari la cui disamina deve essere integrata con i dati oggettivi espressi dalle misurazioni di laboratorio. Soltanto un costante aggiornamento e l’esperienza permettono di integrare le opportune valutazioni dell’operatore ad un caso reale. Le caratteristiche genetiche e l’adattamento ecologico delle singole specie possono influire sui ritmi di sviluppo, risultando quindi più appropriato utilizzare i dati in letteratura che si riferiscono a sperimentazioni condotte nella stessa area zoogeografica dei campioni in studio (J. Amendt et al., 2006).
Lo studio delle strutture microscopiche delle larve, come l’apparato boccale e gli “spiracoli” posteriori, è effettuato attraverso sofisticati microscopi in grado di differenziare minuzie morfologiche invisibili ad occhio nudo.
Il ritmo di crescita degli stadi immaturi dei Ditteri dipende essenzialmente dalla temperatura ambientale, quindi è possibile far risalire all’età dei campioni partendo dalla cosiddetta “storia termica” degli insetti, cioè con la somma delle quantità di calore accumulata dagli esemplari durante la loro crescita.
Se lo specialista valutasse il P.M.I. basandosi esclusivamente sulla temperatura media, riferita all’intera durata del ciclo di sviluppo larvale, rischierebbe di dare un’erronea valutazione tanatocronologica. Oltre alle informazioni sulla temperatura ambientale è fondamentale considerare gli ulteriori motivi di elevazione termica, tra cui la presenza e la densità della massa larvale, le quali influenzano la “somma termale” ossia la quantità di calore assorbita dagli insetti. A causa dei movimenti frenetici degli insetti, ad esempio, la temperatura all’interno della massa larvale può essere significativamente più alta rispetto temperatura ambientale (fig. 15) (Wells & LaMotte, 1995); la velocità di sviluppo delle larve è quindi alterata ovvero accelerata dall’energia cinetica dovuta all’azione demolitiva degli insetti. Sono riportati casi in cui all’interno della carcassa la temperatura raggiungeva picchi di 49°C a fronte di una temperatura esterna che oscillava tra i 9 e i 22°C, ed esperienze in cui la massa larvale risultava attiva all’interno del substrato cadaverico nonostante si registrasse una temperatura esterna di -4°C (C.C. Deonier, 1940). Il calore sviluppato dalla massa larvale ha quindi importanti risvolti medico-legali perché in alcune circostanze (ampia massa larvale estesa in più zone del corpo) è in grado di vanificare l’effetto refrigerante prodotto dalla cella frigorifera dove si trova il cadavere.
Uno dei maggiori limiti di questo strumento dipende dal fatto che, sebbene gli insetti vengano attratti dall’odore della carcassa entro pochi minuti dalla comparsa di questa nell’ambiente, ciò non è sufficiente alla deposizione delle uova se la temperatura e le condizioni del substrato non sono ideali. L’ovodeposizione non avviene fin quando non si instaura, complessivamente, quell’insieme di condizioni del substrato (ossia dei tessuti della carcassa) tale da favorire i primi stadi di sviluppo dell’uovo e/o della larva. Le femmine dei Ditteri ad esempio non depongono su tessuti mummificati ma solo su quelli umidi perché solo in queste condizioni le larve possono svilupparsi.
Le successioni: il cadavere, specialmente se esposto all’ambiente esterno, può essere considerato come un ecosistema a sé stante, soggetto a rapidi cambiamenti naturali dovuti alle diverse fasi della decomposizione (diminuzione della temperatura corporea, irrigidimento delle fibre muscolari, autolisi, acidità, etc.) e alla presenza della fauna necrobionte.

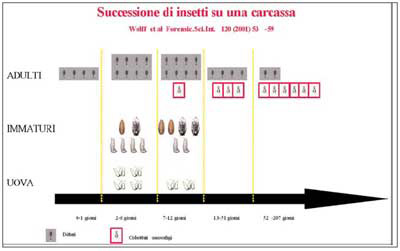
Lo studio delle successioni (successional waves, “successioni di ondate” o “ondate di insetti”) prende in considerazione tutti gli insetti contemporaneamente presenti su un cadavere (fig. 16), i rapporti che ciascuna specie contrae con le altre e il ruolo che i diversi organismi hanno nel modificare il substrato.
Le specie che possono essere raccolte differiscono in base alla regione in cui si lavora ed alle diverse circostanze in cui è avvenuta la morte, ma in linea di massima l’andamento della loro permanenza segue un decorso simile che è stato rappresentato da diversi autori con tabelle in cui sono descritte più “ondate” di colonizzazione in relazione alle fasi di decomposizione del cadavere (Smith, 1986; Turner, 1991). Attraverso questi studi, portati avanti grazie alle sperimentazioni su carcasse animali, in particolare suini, è stato possibile provare che in ogni stadio prevalgono numericamente alcune specie anche se le altri esemplari caratteristici degli altri stadi di decomposizione sono comunque presenti.
Le osservazioni sperimentali dimostrano che i diversi periodi di decomposizione e le particolari successioni di insetti non possono essere interpretati come una sequenza di fenomeni separati l’uno dall’altro, ma devono essere intesi come eventi che si sovrappongono e si combinano tra loro in una successione ininterrotta (fig. 17): questa sovrapposizione rappresenta la principale difficoltà nell’analisi ai fini medico-legali e richiede una provata esperienza da parte dell’operatore nell’interpretare i dati entomologici alla luce degli eventi ambientali.
Molte famiglie di insetti che appaiono presto sulla scena di morte, possono comprendere specie non prettamente necrofaghe (Amendt J et al., 2004); queste specie possono determinare cambiamenti nella microfauna cadaverica attraverso fenomeni di predazione e parassitismo, la comprensione dei quali è indispensabile per non commettere errori interpretativi grossolani.

e. Correlazione campioni entomologici-dati ambientali e calcolo del P.M.I.
Il calcolo del P.M.I. nei casi studiati è basato fondamentalmente sulla correlazione tra lo stadio di vita dell’insetto identificato dopo il campionamento e le temperature ambientali alle quali è stato sottoposto dalla deposizione fino al suo ritrovamento. Questi dati sono confrontati con apposite “tabelle di crescita” messe a punto da vari autori e che prendono in considerazione, per ogni specie di insetto, la tempistica del loro ciclo di sviluppo.
Viene scelta la tabella con dati che meno si distaccano dalla situazione che si vuole studiare e si calcola il P.M.I., che può essere espresso come ADH o ADD(10).
In condizioni normali le femmine dei Ditteri trovano il cadavere entro pochi minuti o poche ore dalla morte ed effettuano subito la deposizione delle uova: in questo modo si attiva immediatamente l’orologio biologico che permette il calcolo del P.M.I. e in particolare si può risalire al “P.M.I. massimo”, periodo di tempo compreso tra la morte con il contemporaneo arrivo degli insetti ed il momento del ritrovamento del cadavere. In particolari circostanze (es. corpo mantenuto in ambiente ermetico, corpo sotterrato o temperature ambientali troppo basse, etc.), però, l’accesso al cadavere da parte dei Ditteri non è possibile o possibile solo dopo un certo periodo: in questi casi l’orologio biologico per il calcolo del P.M.I. non si attiva o si attiva in ritardo, quindi la stima riguarderà solo un parametro più ridotto definito M.P.M.I. o Minimun Post Mortem Interval (P.M.I. minimo), periodo di tempo compreso tra l’arrivo degli insetti e il ritrovamento del cadavere, senza però ottenere informazioni sull’effettiva data di morte.
L’Italia, in considerazione della sua particolarissima connotazione geografica, esprime condizioni ambientali molto varie secondo la latitudine, l’altitudine, la vicinanza del mare e/o di bacini idrici, la stagione e le specie che in un dato momento ed in un dato microclima hanno trovato le condizioni ottimali di sviluppo. Se a ciò si aggiunge che le specie di insetti necrofagi rinvenibili sul luogo del delitto oltre a essere quantitativamente numerose presentano ognuna diversi stadi di maturazione, tutti diversamente influenzabili dall’ambiente, si capisce quanto sia importante la profonda conoscenza degli animali in questione ed il loro comportamento in esperimenti controllati ed in casi reali.
4. Sopralluogo, campionamento e conservazione dei campioni entomologici
Gli insetti che si trovano su un cadavere o vicino ad esso, se correttamente raccolti, conservati e analizzati da un entomologo forense esperto e messi a confronto con dati ambientali rilevati in maniera accurata, possono fornire stime oggettive del momento della morte e offrire altre valide informazioni relative alle circostanze del decesso.
Sono state descritti in letteratura numerosi sistemi di raccolta/cattura, nelle più svariate scene di morte (Lord & Burger, 1983; Goff, 1998b; Haskell & Williams, 1990; Haskell et al., 1997), che prevedono accorgimenti generali, ma nella pratica ogni entomologo perfeziona il proprio metodo grazie all’esperienza acquisita e maturata attraverso il confronto con gli altri operatori del settore.
Per lo svolgimento di tali attività sono state ideate delle schede (Amendt et al., 2006; Magni et al., in stampa), da compilare in sede di sopralluogo e di autopsia, che possono aiutare il medico legale e le Forze dell’Ordine e l’entomologo a reperire il maggior numero di informazioni (biologiche, climatiche e circostanziali) utilizzabili per il successivo compimento delle indagini delegate.
Le finalità della presente trattazione non consentono di sviscerare nel dettaglio tali aspetti, ma è utile porre l’accento su alcuni concetti base che, se applicati, possono contribuire ad evitare la perdita delle preziose informazioni che l’entomofauna necrobionte può fornire.
Per ciò che attiene al sopralluogo, come per le tecniche ispettive classiche, vige l’assioma “Initial Success or Final Failure”(11). Tali espressioni suggeriscono l’influenza che possiedono il sopralluogo e la strategia d’osservazione e repertamento sulla scena criminis nella estrapolazione di dati informativi. In tale fase è estremamente importante documentare tutte le azioni intraprese e, tramite l’ausilio di mezzi video-fotografici, i siti interessati da colonizzazione della microfauna e l’ambiente circostante. La ricerca di tali siti va estesa alle zone immediatamente adiacenti il cadavere (sotto il corpo, tra le coperte, dietro la tappezzeria, cavità dei muri, condotte di areazione, etc.) ma anche alle zone distanti alcuni metri; ciò può essere indispensabile per il ritrovamento degli esemplari negli stadi successivi a quello larvale. Tali siti vengono scelti dagli insetti stessi in base alle condizioni ottimali per lo sviluppo successivo (Larva Pupa Adulto). Una volta terminato il proprio ciclo di crescita, gli adulti, possono ulteriormente “modificare” la scena criminis trasportando tracce di sangue della vittima anche in luoghi distanti, complicando quindi l’analisi delle macchie di sangue o “B.P.A.” (Benecke et al., 2003).
è molto importante descrivere dettagliatamente le condizioni ambientali al momento dei rilievi tecnici nonché rilevare quelle dei giorni passati e quelle dell’intervento dei primi soccorritori (Forze dell’Ordine, Vigili del Fuoco, etc.). A tal proposito bisogna evidenziare che, nel caso di luoghi chiusi, l’errata abitudine di aerare i locali mediante l’apertura delle finestre e delle vie d’accesso al fine di allontanare l’odore pungente, altera irrimediabilmente il succitato binomio “entomofauna-ambiente”. L’entrata di “nuovi” insetti, la brusca variazione della temperatura e le modificazioni che entrambe possono produrre sulla scena introducono un imponderabile fattore d’incertezza nelle valutazioni entomologiche. Durante il sopralluogo l’Entomologo Forense deve necessariamente interagire con gli altri operatori sulla scena criminis al fine di adottare le più opportune strategie comuni di rilievi e repertazione, senza ostacolare (o peggio alterare) il campo d’azione delle altre tipologie di analisi forensi. Risulta qui evidente che gli interventi sulla scena del crimine debbano avere una completezza multidisciplinare e che debbano essere necessariamente espletati da persone preparate e qualificate ossia da specialisti in grado di recuperare e conservare le prove riguardanti le diverse discipline (Magni et al., in stampa). Solo adottando tale strategia le tracce, analizzate ognuna dal singolo professionista e correlate tra loro da un team di specialisti, potranno fornire indicazioni riguardo ai periodi ante-mortem, peri-mortem e post-mortem, ricostruendo gli eventi significativi di un crimine.
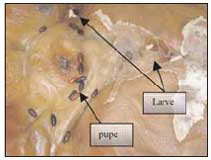
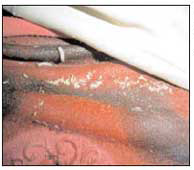
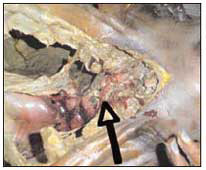
Il Campionamento - È importante raccogliere campioni rappresentativi di tutti gli adulti e gli stadi immaturi che si trovano sopra, sotto ed attorno al cadavere. Il campionamento prosegue anche durante l’autopsia, importante occasione per controllare accuratamente le pieghe dei vestiti, tipica zona d’annidamento delle pupe (fig.18) e per reperire materiale all’interno del corpo (ferite, pieghe cutaneee, cavità cranica, canali digestivi e respiratori se colonizzati possono favorire lo sviluppo di una fauna diversa che nell’ambiente circostante) (fig. 19-20-21).
La raccolta degli insetti adulti può avvenire con diversi sistemi (rete aerea, trappole appiccicose, etc.) ma per ciò che attiene alle larve e alle uova, l’uso rispettivamente di cucchiaio e pennellino è strettamente necessario per non danneggiare le microstrutture altamente identificative. Sulla scena del ritrovamento dopo la rimozione del corpo, è indispensabile inoltre prelevare campioni di foglie e di terreno in quanto questi luoghi sono il posto ideale in cui gli insetti si lasciano cadere o si nascondono per ripararsi ed “impuparsi”.
Generalmente i campioni sono suddivisi in due sottocampioni: un gruppo è ucciso per fermare l’orologio biologico ed un gruppo è mantenuto in vita ed allevato per confermare l’identificazione della specie con l’analisi dell’insetto adulto (Lord & Burger, 1983; Haskell & Williams, 1990).
La Conservazione - La conservazione e la collezione degli insetti segue procedimenti diversi in base al tipo di campione da mantenere: solamente gli esemplari adulti vengono spillati, mentre gli stadi giovanili rimangono sotto alcool all’interno di provette sigillate. Il punto essenziale rimane però il trattamento immediato delle uova e delle larve al fine di conservarne lo stato di ritrovamento. Tale accorgimento preclude la possibilità per gli insetti di crescere (o decrescere) ed eventualmente di andare incontro al successivo stadio del ciclo vitale, fenomeni che falserebbero in modo notevole la stima del P.M.I. è fondamentale che le larve campionate vengano poste in acqua bollente per pochi secondi e in seguito conservate in alcool etilico al 70%: il processo di bollitura permette di mantenere quasi totalmente inalterate le dimensioni dei campioni, che possono essere così misurati senza incorrere in errori grossolani, dovuti all’effetto disidratante delle soluzioni conservanti (le soluzioni disidratanti causano un accorciamento “meccanico”). è comunque consigliabile l’identificazione e la misurazione del campione il prima possibile dopo la conservazione (Amendt et al., 2006). In generale le metodologie di campionamento e conservazione possono essere modificate leggermente secondo il contesto operativo ma è solo l’esperienza dell’operatore a guidare la scelta verso la soluzione più opportuna.


5. Il contributo della biologia molecolare all’entomologia forense
Nell’ultimo ventennio l’avvento delle nuove tecniche della biologia molecolare ha rivoluzionato il mondo della medicina, della microbiologia, delle biotecnologie ed ovviamente del settore forense aumentando esponenzialmente il volume di conoscenze in nostro possesso. Molte delle informazioni riguardanti il patrimonio genetico, il quale rappresenta al tempo stesso l’origine della biodiversità ed il depositario dei caratteri ereditari, sono state acquisite grazie ad un rapido ed progressivo avanzamento delle frontiere della biologia molecolare moderna. Se da un lato ciò ha contribuito ad inasprire la polemica sull’uso indiscriminato di tali strumenti a fini non direttamente terapeutici (es. clonazione, fecondazione artificiale, OGM, etc.), dall’altro lato tali scoperte hanno apportato innovazioni straordinarie in molti campi delle scienze applicate tra cui quello della biologia forense. In particolar modo, al fine di soddisfare la richiesta di metodi affidabili, sensibili e altamente discriminanti, l’analisi del DNA per scopi identificativi(12) ha subito una eccezionale propulsione.
Le metodologie in uso, ormai validate a livello mondiale, permettono oggi un potere discriminante dell’ordine di 1 su 1016 individui (in altre parole 1 su 10.000.000.000.000.000). La sensibilità di tali tecniche, cioè la soglia minima necessaria alla rilevazione del materiale genetico, si è abbassata notevolmente, passando in pochi anni da un valore di 100 nanogrammi (cioè miliardesimi di grammo) di DNA genomico al limite attuale di poche decine di picogrammi (cioè kilomiliardesimi di grammo) raggiunto grazie alla messa a punto di protocolli di estrazione specifici per il DNA in LCN(13) (Saravo et al., 2003) ed all’ausilio di sofisticate tecniche di micro-dissezione laser (Di Martino et al., 2006, Staiti et al., 2005). La scoperta di nuovi “marcatori”(14), infine, sta incrementando enormemente l’arsenale di strumenti a disposizione dei biologi forensi, non solo elevando il potere di discriminazione nei casi controversi (Di Luise et al., in press) ma anche aumentando la percentuale di successo nell’analisi di reperti antichi e di campioni altamente degradati (MiniSTR, SNiPs) (Romano et al., 2006) ovvero a discernere i contributi maschili da quelli femminili in una miscela biologica come nei casi di violenze sessuali (marcatori dei cromosomi X e Y). La stessa biologia molecolare che ha permesso i suddetti traguardi, può contribuire attivamente alle analisi dell’entomologia forense. In particolare possiamo distinguere due linee di applicazione della biologia molecolare all’ Entomologia Forense:
a. Identificazione mediante analisi molecolare delle specie rinvenute sul luogo del reato. L’utilizzo delle tecniche di biologia molecolare è in questo caso finalizzato alla precisa identificazione di una specie di insetto. Quest’ultima, infatti, se basata sulla metodologia classica, può presentare dei grossi limiti. L’approccio classico è basato sull’individuazione di caratteri morfologici che possono essere ritrovati nelle cosiddette “chiavi dicotomiche”, strumenti indispensabili per l’attribuzione di un esemplare ad una particolare specie. Il limite di questo approccio è che per alcune specie non è però sempre agevole o addirittura possibile distinguere morfologicamente le uova, le pupe e gli stadi larvali dagli stadi analoghi di specie diverse (Amendt, et al., 2004). Inoltre tale approccio può diventare molto laborioso e lungo, potendo richiedere anche l’allevamento (per diverse settimane e non sempre con esito positivo) degli esemplari campionati al fine di esaminarne gli stadi di sviluppo più avanzati. Una conoscenza analitica, sistematica e completa di grandi Ordini di insetti è ottenibile solo dopo numerosi anni di studio ed applicazione pratica ed è appannaggio solo di poche nicchie di ricercatori. Infine la valutazione che si ottiene dall’applicazione di tali misurazioni è in ogni caso ombreggiata da un velo di soggettività che potrebbe in alcuni casi pregiudicare una piena “efficacia dibattimentale”. Con l’avvento delle tecniche basate sull’utilizzo del DNA sono state identificate alcune regioni del genoma comuni agli appartenenti alle stesse specie e/o a specie affini; si è così scoperto che alcune di queste potevano essere considerate dei veri e propri “contrassegni” molecolari o “marcatori”, cioè sequenze in grado di identificare e classificare univocamente una specie. L’entomologia classica e la zoologia in generale hanno visto rivoluzionare il proprio campo di studio dall’avvento di tale metodologia di classificazione, ma al tempo stesso hanno potuto testarne le potenzialità ed apprezzarne i vantaggi. Nei primi anni di applicazione dei marcatori molecolari ai fini tassonomici(15) molte specie animali che prima erano state classificate, tramite l’approccio delle chiavi dicotomiche, ad un particolare taxon, vennero in seguito attribuite a gruppi filogeneticamente molto distanti, contribuendo ad incastrare i tasselli mancanti nello studio dell’evoluzione: si passò così da chiavi “fenotipiche” a chiavi “genotipiche”. Numerosi studi sono stati pubblicati sull’identificazione per via biomolecolare degli invertebrati di interesse medico-legale. Alcuni di essi sono il risultato di un applicazione pratica a casi reali in cui l’approccio classico non era stato soddisfacente. Non solo le larve e gli adulti campionati sul luogo del reato ma anche i “pupari” (gli involucri protettivi dalla larva in metamorfosi) si sono dimostrati utili ai fini identificativi, sebbene fosse trascorso un certo intervallo di tempo dalla scomparsa degli esemplari che li avevano prodotti (cioè degli esemplari presenti nelle prime fasi della colonizzazione) (Mazzanti et al., 2006). Il grande vantaggio dei dati ottenuti grazie al sequenziamento del DNA è quello di applicarli in maniera speditiva e, una volta informatizzati grazie all’ausilio di moderni software, interpretati alla luce di calcoli statistici complessi (Sperling et al., 1994). In generale si possono distinguere due oggetti dell’analisi biomolecolare utilizzati per l’identificazione di specie: marcatori del DNA mitocondriale (mtDNA) e marcatori del DNA nucleare. Nel primo caso i marcatori maggiormente utilizzati sono le sequenze nucleotidiche che codificano per le subunità I e II (COI e COII) delle citocromo ossidasi, proteine enzimatiche coinvolte nel metabolismo energetico. Sono inoltre utilizzate, anche per altri Ordini, le sequenze delle subunità 12S e 16S del RNA ribosomiale, il gene del tRNA della leucina. Tali sequenze sono messe a confronto con le sequenze di riferimento ottenibili in apposite banche dati su internet o presso musei di scienze naturali in tutto il mondo. La perfetta sovrapponibilità delle sequenze è sufficiente, insieme con i dati morfo-fisiologici, per un’accurata identificazione di specie. Differenze della sequenza nucleotidica possono invece essere attribuite a variabilità intraspecifica (tra esemplari della stessa specie) o interspecifica (tra specie dello stesso genere e di generi diversi). Con numerosi esperimenti condotti su esemplari di riferimento, gli studiosi Wells, Sperling e Vincent hanno dimostrato che la variabilità intraspecifica delle sequenze COI e COII è comunque minore o uguale all’1% mentre quella interspecifica si attesta intorno al 3-5% (Wells e Sperling, 2000-2001; Vincent, 2000). Sebbene non si possa escludere a priori un fenomeno di sovrapposizione delle due tipologie di variabilità, questi dati fissano una linea di demarcazione tra la percentuale di differenze nucleotidiche presenti all’interno di una data specie e l’analoga percentuale riscontrabile nel confronto di specie differenti.
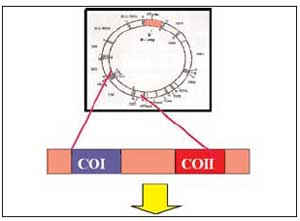
In pratica il rilevamento di variazioni di un ordine minore o uguale all’1% nella sequenze COI e COII del DNA mitocondriale potrà essere assunto come effetto della naturale variabilità genetica tra individui simili, mentre percentuali superiori al 3% saranno attribuiti alla presenza di specie differenti. L’approccio biomolecolare si dimostra quindi adeguato per l’identificazione a livello di specie anche alla luce del fatto che non sono state osservate variazioni intraspecifiche significative tra esemplari campionati in regioni molto distanti del globo terrestre (Stevens and Wall, 1996, 2001). Non bisogna altresì dimenticare che la ricerca zoologica per l’identificazione di specie non di interesse forense ha portato negli ultimi anni alla scoperta di nuovi marcatori che in futuro potrebbero risultare utili per il differenziamento di ceppi della stessa specie. Sperling ha inoltre considerato la possibilità che gli studi sulla sequenza delle citocromo ossidasi, proteine fondamentali per il metabolismo, possano contribuire allo sviluppo di sistemi predittivi circa l’accrescimento e la durata degli stadi larvali (Sperling et al., 1994). Infine è doveroso segnalare che, a fronte delle innegabili potenzialità di tali tecniche, sono stati riportati in letteratura alcuni casi d’errata identificazione di specie a causa di ibridazione interspecifica (accoppiamento tra specie dello stesso genere), fenomeno peraltro non raro negli artropodi (Stevens et al., 2002). L’utilizzo di marcatori del DNA nucleare prevede invece l’utilizzo delle regioni ITSI e ITSII, lsu-28S, D1-D7. In particolare l’analisi della subunità grande (large subunit) del RNA ribosomiale 28S si è rilevata molto sensibile nell’identificazione e discriminazione tra specie appartenenti alle famiglie delle Calliphoridae e Luciliinae (Stevens and Wall, 2001). L’utilizzo di altri marcatori del DNA nucleari (STR) appare controverso in questo tipo di applicazione essendo stati rinvenuti esemplari della stessa specie (ma di popolazioni molto distanti) che esibivano una variabilità genetica molto accentuata ossia dello stesso ordine di grandezza di quella esistente tra due specie diverse. Infine è importante sottolineare che, mentre è nota la sequenza del mtDNA di almeno 400 specie, quella dei marcatori nucleari e delle regioni adiacenti è ancora poco completa. Recentemente sono stati intrapresi degli studi per valutare la possibilità di identificare univocamente lo stadio di vita attraverso l’espressione del RNA messaggero ed i fattori di controllo della trascrizione che caratterizzano i diversi cicli vitali (Ames et al., 2004);
b. applicazione al fine di accertare la correlazione esistente tra insetto, luogo del reato e vittima. Alcuni rappresentanti dell’entomofauna necrobionte, come prima illustrato, si nutrono dei tessuti del corpo (necrofagi) oltre che dei colonizzatori primari (predatori). Alcuni insetti, detti ematofagi, assumono il sangue dai corpi in quanto essenziale per una completa maturazione delle uova prima della deposizione. Il materiale biologico della carcassa può quindi permanere indigesto all’interno di tali esemplari per un tempo variabile, costituendo una fonte di DNA utilizzabile ai fini investigativi (Introna, 1999; Clery, 2001). Nel caso di spostamento/occultamento del cadavere, il ritrovamento e la localizzazione di particolari insetti, o meglio del loro contenuto di DNA umano, può rappresentare la prova del “passaggio” della vittima in un determinato luogo. Similmente tale approccio può chiarire, in base allo studio delle successioni, dove e/o su cosa si siano sviluppate le larve nel caso in cui la “fonte alimentare” delle stesse sia dubbia. Anche i detriti e le feci prodotte da tali insetti possono contenere quantità di DNA umano sufficienti all’identificazione del cadavere a scapito del quale essi si sono nutriti (Reploghe, 1994). Nel 2002 un gruppo di ricercatori guidati da J. Di Zinno del Laboratory Division del F.B.I. riuscì a sequenziare il DNA mitocondriale contenuto nel tratto intestinale di scarafaggi (coleotteri) che si erano sviluppati su ossa umane.
Alcuni Autori (Campobasso et al. 2005) hanno valutato i parametri fondamentali per l’utilizzabilità del DNA umano contenuto nel “gozzo” (crop), un’appendice esofagea dei Ditteri con funzioni d’immagazzinamento del cibo. In particolare questi ricercatori hanno considerato l’età e le dimensioni ideali, il tempo limite dal campionamento all’analisi molecolare, le metodologie per prevenire le contaminazioni dei tegumenti esterni della larva, i fenomeni degradativi del DNA all’interno del gozzo e l’influenza dei metodi e dei tempi di conservazione. Le conclusioni, sebbene complesse ed articolate, indicano che, entro un range compreso tra 2,5 e 4,5 giorni dal campionamento, le dimensioni del gozzo ed il ciclo vitale della larva rappresentano le variabili più importanti; il tempo di conservazione, la lunghezza complessiva del P.M.I. ed gli eventuali fenomeni degradativi a carico del materiale genetico sembrerebbero avere una scarsa incidenza nella “tipizzazione” del DNA umano (Wells et al, 2001). Recentemente un gruppo di studiosi ha dimostrato la possibilità di rilevare l’antigene prostatico specifico (PSA) e di tipizzare i marcatori del cromosoma Y partendo dall’estratto del “gozzo” di Ditteri allevati parecchie ore prima (fino a 145 ore) su tessuti ricoperti di sperma umano (Clery, 2001). Il test del PSA ed i marcatori genotipici del cromosoma Y sono correntemente utilizzati nei casi di violenza sessuale rispettivamente per determinare la presenza di sperma e per distinguere il contributo maschile nelle miscele genotipiche (genotipo aggressore + genotipo vittima) che si ottengono dall’analisi degli indumenti e/o del tampone vaginale della vittima. L’esperimento di Clery e dei suoi collaboratori apre la strada all’applicazione ai casi di violenza sessuale-omicidio in cui i cadaveri vengono rinvenuti molti giorni dopo l’evento delittuoso. In considerazione del fatto che le vittime sono a volte ritrovate in condizioni tali da non permettere l’isolamento delle tradizionali tracce biologiche dell’aggressore (sul corpo e sui vestiti) e che gli orifizi del cadavere sono i punti più velocemente occupati dall’entomofauna, tale strategia potrebbe essere decisiva nell’identificazione del colpevole. In casi di violenza sessuale è anche possibile il trasferimento occasionale di pidocchi o piattole, parassiti comuni dell’Uomo, dall’assalitore alla vittima; giacché tali artropodi assumono sangue umano e, considerato che è stata verificata la rilevabilità del DNA umano nei loro resti, essi potrebbero essere utilizzati per accertare la correlazione fisica dell’ “ospite originario” con la vittima (Mumcuoglu et al., 2004). Data la peculiare composizione cellulare dell’invertebrato e dell’esigua (e/o degradata) quantità di DNA, è auspicabile comunque l’applicazione di metodiche estrattive modificate e di laboratori attrezzati per l’analisi in alta sensibilità (Spitaleri et al., 2006; Campobasso et al., 2005).
Nei casi di correlazione tra insetto, luogo del reato e vittima, i marcatori molecolari (STR - Short Tandem Repeats) sono gli stessi utilizzati nella routine dai laboratori delle Forze dell’Ordine per l’attribuzione di tracce biologiche repertate sulla scena criminis ad un soggetto sospettato o indagato. I dati ottenuti quindi sono direttamente comparabili con quelli presenti negli archivi dei laboratori delle Forze dell’Ordine o con quelli eventualmente acquisiti nell’ambito delle attività di Polizia Giudiziaria.
6. Cenni di entomotossicologia
L’entomotossicologia studia l’applicazione delle analisi tossicologiche agli insetti cadaverici, al fine di identificare droghe e tossine presenti nei tessuti e per determinare gli effetti causati da queste sostanze sullo sviluppo degli artropodi e quindi sul calcolo del P.M.I. (Introna et al., 2001). Nel caso in cui il cadavere si trovi in uno stadio di decomposizione particolarmente avanzato, i fluidi biologici e gli organi solidi potrebbero non essere più disponibili per le tradizionali analisi tossicologiche. In questi casi gli esemplari di insetti sono utilizzabili per diversi tipi di analisi tossicologiche, soprattutto di carattere qualitativo (Nolte et al., 1992; Goff & Lord, 1994, 2001; Introna et al., 2001; Campobasso et al., 2004; Traqui et al., 2004). Il passaggio dei composti tossici e delle sostanze venefiche dalla carcassa ai ditteri non si esaurisce al primo anello della catena alimentare, ma prosegue anche nei coleotteri, predatori delle larve (Bourel, 1998), in cui si nota un bioaccumulo secondario.
Le larve che si nutrono della carcassa possono accumulare le droghe ed i farmaci di cui aveva fatto uso il soggetto prima di morire; inoltre è possibile rilevare numerose sostanze tossiche alle quali il soggetto era stato esposto in vita. Dopo l’omogenizzazione delle larve sono possibili diversi tipi di analisi come la cromatografia su strato sottile (TLC), il saggio radioimmunoenzimatico (RIA), la gas-cromatografia (GC), la gas-cromatografia/spettrometria di massa (GC/MS) e la high-performance liquid chromatography/spettrometria di massa (HPLC/MS) (Gagliano-Candela and Aventaggiato, 2001; Goff and Lord, 2001); queste tecniche possono essere applicate anche a insetti adulti, frammenti di pupari (Goff et al., 1993, 1997 ; Miller et al., 1994; Introna et al., 1996), esuvie e feci di coleotteri che è possibile rinvenire sulla scena del crimine già dopo i primi giorni ma anche a molti anni di distanza dall’evento criminoso (Nortueva et al., 1982; Miller et al., 1994; Amendt et al., 2004).
Per alcune sostanze i dati tossicologici ottenuti dalle larve di insetti risultano essere più attendibili di quelli ricavati dai tessuti cadaverici; residui di paracetamolo, benzodiazepine, barbiturici e antidepressivi triciclici possono essere rilevate in larve campionate su un cadavere dopo molti giorni dalla morte (Kintz et al., 1990). L’ingestione di droghe o di sostanze tossiche può influenzare lo sviluppo degli insetti necrofagi (O’Brien and Turner, 2004; Pien et al., 2004), come dimostrano esperimenti condotti su carcasse di conigli uccisi con una dose letale di cocaina (Goff et al., 1989) o di eroina (Goff et al., 1991): il ritmo di crescita degli insetti allevati sui fegati e sulle milze di questi animali è molto più rapido che in condizioni standard. Caso contrario avviene con la morfina, che rallenta il ritmo di crescita degli insetti che si nutrono dei tessuti che la contengono (Bourel, 1999).
Tuttavia, come dimostrano gli studi di Sadler et al. (1997), le sostanze stupefacenti si accumulano nell’organismo dei ditteri in maniera non prevedibile, quindi rendendo tale tecnica, sicuramente molto interessante per le finalità di P.G., ancora abbisognevole di ulteriori approfondimenti.
In un interessante studio Roeterdink e collaboratori (2004) hanno dimostrato che le larve di una specie di Calliphora, recuperate da tessuti animali in decomposizione, possono accumulare le particelle di tipo GSR o Gunshot residues, ossia i residui da polvere da sparo comunemente ricercati per stabilire l’utilizzo di armi da fuoco. Ciò può risultare molto utile nel caso in cui l’assenza di ogive e l’avanzato decadimento del cadavere non permettano il riconoscimento delle tipiche ferite da arma da fuoco. In particolare gli esperimenti preliminari di questi studiosi hanno permesso di ipotizzare che le larve necrobionti fungessero da bioaccumulatori del Bario (uno degli elementi chimici presenti nelle GSR Piombo-Antimonio-Bario) durante le prime fasi di accrescimento larvali. Piombo, Antimonio e Bario subiscono comunque un decremento globale in seguito all’allontanamento degli insetti dal tessuto contaminato con tali elementi.
7. Casi reali
In passato le segnalazioni casistiche hanno rappresentato la bibliografia di riferimento dell’entomologia forense ed ancora oggi i molti casi illustrati dagli entomologi e dai patologi contribuiscono alla diffusione degli aspetti più affascinanti di questa disciplina, accrescendo l’interesse degli operatori nel settore investigativo. Sono stati, pertanto, selezionati alcuni casi giudiziari in cui l’aspetto entomologico è stato un prezioso strumento determinante per la risoluzione e che mostrano l’aspetto pragmatico di quanto finora esposto.
1) Influenza dell’esposizione e della copertura della carcassa sul calcolo del P.M.I.
Caso 1. Il corpo nudo di una giovane donna venne scoperto il 31 agosto di qualche anno fa in una scarpata nei pressi di una strada poco distante dall’aeroporto di Seattle (USA). Il cadavere era esposto al sole e la parte inferiore del corpo si trovava all’interno di un sacco di plastica nera per rifiuti. Uova di mosca furono campionate dalla metà superiore del corpo; la raccolta fu eseguita anche per le numerose larve che infestavano le aperture vaginale, orale e nasale. I dati ambientali ottenuti dalla stazione meteorologica dell’aeroporto registravano temperature massime variabili tra 23°C e 33°C, mentre la temperatura del corpo era di 31°C. Tali elementi, se meramente applicati alle tabelle di crescita, avrebbero portato alla stima di un P.M.I. certamente superiore ai tre giorni. Ma, dopo l’allevamento delle larve ed un’analisi dettagliata delle condizioni di sviluppo, gli entomologi conclusero che la vittima era stata uccisa 2,5-3 giorni prima della scoperta del cadavere. Le alte temperature, l’esposizione continua alla luce diretta del sole e la circostanza che la parte inferiore del corpo fosse coperta da un sacco di plastica di colore scuro hanno determinato una crescita delle larve molto più rapida che in condizioni normali. Sviluppi investigativi consentirono di accertare che la vittima era deceduta, per cause accidentali, la notte del 28 agosto (Catts & Haskell, 1990).
Caso 2. I resti carbonizzati di un uomo di 25 anni furono rinvenuti nel mese di agosto all’interno di un’automobile bruciata, in un’area rurale della città di Brindisi (Introna et al., 1998). I resti erano per lo più combusti il che rendeva la determinazione del P.M.I. praticamente impossibile. Tuttavia una parte del cervello e degli organi interni, meno devastati dalle fiamme, presentavano colonizzazione da parte di larve di Ditteri. L’autopsia rivelò due fori da proiettile nella zona del collo e contenuti stomacali parzialmente digeriti costituiti da carne e sostanze vegetali. L’uomo fu riconosciuto grazie al confronto delle impronte dentarie e, sulla base delle specie di insetti rinvenuti sulla scena del crimine, il P.M.I. venne stimato in 3-4 giorni. Approfondimenti investigativi confermarono tale dato in quanto l’ultima volta in cui l’uomo era stato visto in vita risaliva proprio a quattro giorni prima del rinvenimento del corpo quando, insieme con un gruppo di amici, si recò in un ristorante e ordinò carne e insalata (Introna et al., 1998).
2) Rilevanza dell’ambiente e delle azioni antropiche sullo sviluppo della biocenosi cadaverica.
I corpi di due donne vennero rinvenuti in ottobre, a distanza di pochi giorni l’uno dall’altro, in due campi di grano limitrofi situati sulla terraferma in prossimità della città di Venezia. Entrambi i cadaveri mostravano chiare lesioni da strangolamento. La somiglianza tra i due casi, sia geografica sia temporale, avrebbe fatto supporre una simile colonizzazione da parte della fauna cadaverica, ma l’analisi dei dati entomologici non confermò questa ipotesi. In entrambi i casi fu campionato Stearibia nigriceps, dittero tipico dello stadio di fermentazione “casearia” che avviene 3-6 mesi dopo la morte: la colonizzazione da parte di questo Piofilide è facilmente spiegabile data l’accelerazione della decomposizione dovuta alle alte temperature estive. Ma solo in uno dei due casi, invece, fu riscontrata la presenza di Hermetia illucens, dittero che aveva inibito la presenza delle normali mosche cadaveriche: infatti, in questo caso, l’entomofauna cadaverica non comprendeva i Calliforidi che invece erano abbondantemente presenti sull’altro cadavere (Turchetto et al., 2001). Tale diversità avrebbe potuto screditare eventuali correlazioni temporali tra i due eventi delittuosi. Ulteriori indagini sul caso permisero di appurare che nei due diversi campi di grano veniva fatto uso di pesticidi di diverso tipo: l’assenza dei Calliforidi causata dalla competizione di Hermetia illucens sul cadavere rinvenuto in una zona più “inquinata” è giustificata dalla maggiore resistenza di tale dittero all’esposizione agli insetticidi (Vanin, 2006). In considerazione di queste valutazioni e di quelle strettamente anatomo-patologiche è stato possibile ricondurre il periodo di morte per entrambi i casi a due mesi prima del ritrovamento.
3) Presenza di fauna cadaverica non specializzata.
I resti scheletrizzati di una giovane donna furono trovati il 29 gennaio in una foresta di querce e aceri delle montagne del Cumberland (USA). Nei pressi del corpo vennero ritrovati un paio di jeans usurati e fu rinvenuto un cavo elettrico, probabilmente utilizzato per strangolare la vittima. L’interno del cranio era occupato da un nido di vespe (gen. Polistes), costituito da circa 100 celle, quasi tutte vuote: questo indicava che una colonia aveva costruito e occupato il nido l’estate dell’anno precedente. Basandosi sulla conoscenza del comportamento delle Polistes è stato possibile supporre che il cranio era libero da tessuto già in primavera, quando le vespe vanno in cerca di un luogo fresco e protetto dove costruire il favo.
All’interno del cranio venne inoltre ritrovato un pupario di Sphaeroceridae, mosca della “seconda ondata”, che permise di indicare che la successione della fauna necrobionte avvenne durante la stagione calda precedente alla costruzione del nido, quindi un intervallo post-mortem di 18 mesi. Questa conclusione fu sostenuta in modo indipendente da un antropologo forense in base ai segni delle intemperie sullo scheletro e da specialisti che analizzarono e datarono i segni d’usura del pantalone rinvenuto. L’identificazione della donna, avvenuta grazie alle radiografie dentarie, permise di sapere che la sua scomparsa era stata segnalata il febbraio di due anni prima (Catts & Haskell, 1990).
4) Entomologia Forense e Botanica Forense: l’incidenza di un approccio multidisciplinare alle indagini tecnico-scientifiche.
Caso 1. Nei sobborghi di Chicago una donna venne brutalmente violentata da un uomo che indossava una mascherina da sciatore. Gli investigatori arrivarono un anno dopo ad un sospettato e, muniti di mandato di perquisizione, trovarono nella sua casa una maschera simile a quella descritta dalla vittima dell’aggressione. Interrogato sulla maschera l’uomo ammise la proprietà della maschera, ma affermò di non averne fatto uso dall’ultimo inverno.
Gli investigatori osservarono che alla maschera aderivano molti residui vegetali, tra cui parecchi frutti di Xanthium (frutti aculeati). Nel tentativo di identificare tale reperto per altri fini, gli analisti rilevarono la presenza di piccoli bruchi che vennero successivamente inviati ad un entomologo per l’identificazione di specie. Tali bruchi hanno un ciclo di sviluppo lungo un anno: in primavera gli adulti sono attivi ed all’inizio dell’estate depongono uova da cui, verso la seconda metà dell’estate, si originano bruchi che colonizzano i frutti. Gli stessi bruchi entrano nello stadio di pupazione durante l’inverno ed emergono come adulti fecondi la primavera successiva. L’entomologo concluse che la maschera doveva essere esposta in ambiente esterno durante l’estate precedente, lo stesso periodo in cui avvenne la violenza, e che verosimilmente in quel periodo i frutti di Xanthium erano rimasti impigliati alla maschera, portando al loro interno i bruchi. Quando gli furono contestate queste evidenze scientifiche, il sospettato confessò la violenza (Catts & Haskell, 1990).
Caso 2. In Sicilia il corpo esanime di una prostituta fu trovato tra i cespugli presenti nell’area adiacente ad una spiaggia. L’autopsia rilevò evidenti segni di strangolamento ma nessuna traccia utilizzabile ai fini dell’identificazione dell’ autore del reato. Qualche tempo dopo, le indagini degli investigatori concentrarono i sospetti su un distinto uomo d’affari residente in una provincia dell’entroterra, lontana decine di chilometri dalla spiaggia. Il sopralluogo nell’abitazione del sospettato sembrava essere destinato ad un fallimento investigativo poiché la casa appariva caratterizzata da un perfetto ordine e da una maniacale pulizia. Agli esperti della scena criminis non sfuggì però, su una delle pareti, una zanzara (Culex pipiens) schiacciata ed interessata da una piccolissima macchia rossastra. Alcuni frammenti fogliari presenti sui vestiti ed un vecchio paio di scarpe furono sequestrati per le successive analisi. L’analisi sul DNA, espletata secondo metodologie tali da ridurre i fenomeni di inibizione e da incrementare la soglia di sensibilità, stabilì che il sangue assorbito dalla zanzara apparteneva alla vittima. Il terreno risultò essere sabbia della stessa tipologia del luogo del ritrovamento, mentre i frammenti fogliari, sottoposti ad esame morfologico comparativo da un laboratorio universitario, risultarono appartenere alla Calendula Marittima, una pianta cespugliosa i cui rami nascondevano parzialmente il corpo della vittima al momento del ritrovamento. Se da un lato la sabbia era comune solo ad alcune spiagge lungo la costa, l’associazione di tale sedimento con la Calendula marittima, una pianta alofila di per sé molto rara, poteva considerarsi unico nell’intera isola. è qui fondamentale evidenziare che nessuno di questi risultati, da solo, avrebbe provato una correlazione fisica tra la vittima ed il sospettato ma la bassissima probabilità del verificarsi congiunto di tutti e tre gli eventi, insieme ad altre prove indiziarie, condusse il tribunale ad una condanna per omicidio preterintenzionale (Spitaleri et al., 2006).
5) Variazioni indotte da condizioni climatiche estreme.
Caso 1. A metà del mese di dicembre due sciatori non fecero rientro a casa. Dopo una settimana i soccorsi abbandonarono le ricerche. A metà del mese di maggio ai piedi di una scarpata venne notata una cavità dove fu possibile distinguere un cadavere. Furono catturate alcune mosche che si trovavano nella zona del ritrovamento del corpo. Le registrazioni fatte sul posto indicavano una temperatura di 2,5 e 2,9°C nelle immediate vicinanze del cadavere. Furono campionati insetti vicino agli occhi, sulla parte posteriore del collo e dietro l’orecchio sinistro, vennero rinvenute anche uova, poste in seguito in una teca per allevamento. I campioni vennero attribuiti alle specie Calliphora vicina e Calliphora vomitoria. La presenza di queste mosche in un ambiente estremo, caratterizzato dalle rigide temperature d’alta montagna, conferma la capacità d’adattamento e l’ubiquità di questi animali che è spesso causata dall’insediamento antropico; gli autori ipotizzano che questi insetti siano attirati, oltre che dai cadaveri in stadio fresco, anche da quelli vecchi di anni, ma conservati in modo ottimale dalla neve e dal ghiaccio (Wyss et al., 2003).
Caso 2. Il corpo di un uomo di 77 anni venne trovato 18 giorni dopo l’ultima volta che era stato visto in vita, in una grotta profonda 10 metri, in una foresta della Svizzera a 1260 metri sul livello del mare. Il cadavere era supino, vestito, senza particolari segni di ferite. La grotta era totalmente buia e la temperatura era costante, di 5°C. Il corpo fu trasportato all’Istituto di Medicina Forense e conservato per tre giorni in camera fredda a 4°C. L’esame autoptico rilevò un ematoma epidurale procurato dalla caduta come causa della morte, ma non il P.M.I. Vennero quindi campionate uova di Ditteri sulla testa e sulla bocca, che messe in allevamento dopo poche ore schiusero e furono identificate come Calliphora vicina.
Al fine di dimostrare che Calliphora vicina è in grado di volare e deporre uova all’interno della grotta e per stimare in modo accurato il P.M.I., gli studiosi depositarono 1.5 kg di carne fresca e 1 kg di fegato di maiale all’interno della stessa grotta, nel punto ove il cadavere era stato rinvenuto. Ogni giorno veniva controllata la presenza di uova le prime delle quali vennero deposte dodici giorni dopo l’inizio dell’esperimento; le uova si schiusero il 21° giorno confermando le informazioni riguardanti il giorno della scomparsa (18 giorni dalla sparizione dell’uomo al ritrovamento, più tre giorni di camera fredda). L’identificazione degli adulti permise di appurare che anche in questo caso si trattava di individui appartenenti a Calliphora vicina, una specie evidentemente favorita in tali condizioni.
Questo caso fornisce importanti conferme circa la capacità di sviluppo di una mosca tipica della decomposizione in condizioni estreme sia di temperatura che di illuminazione (Faucherre et al., 1999).
Caso 3. Le salme di quattro aviatori italiani, partiti in volo il 28 febbraio e dispersi a causa di un incidente aereo, vennero rinvenute nel deserto egiziano il giorno 8 giugno dello stesso anno. I corpi erano ancora parzialmente vestiti e completamente scheletrizzati. I resti furono sottoposti ai rilievi per l’identificazione, per la causa e l’epoca di morte. Il medico incaricato fu colpito in modo particolare dalla grande presenza, fra le pochissime parti molli dei cadaveri, di coleotteri della specie Dermestes frischi (larve, involucri larvali e adulti). I ditteri erano rappresentati solo da pupari vuoti di due specie di Muscidae. Un successivo sopralluogo nella località di ritrovamento delle salme permise di confermare che nessun altro insetto aveva contribuito alla degradazione attiva dei corpi degli aviatori.
Allo scopo di stabilire l’arco temporale in cui si completa la riduzione scheletrica di un corpo posto all’aperto nelle condizioni ambientali tipiche del deserto, vennero poste due carogne di cane protette da una fitta rete metallica per evitare possibili attacchi da parte di volatili e mammiferi carnivori. Tale sperimentazione confermò sostanzialmente i dati ottenuti dal repertamento sulle salme e permise di appurare che in una zona molto calda e secca come il deserto, i fenomeni cadaverici dissolvitori e trasformativi seguono un andamento che si discosta notevolmente da quello descritto in condizioni normali. L’azione della fauna entomologica in questo caso fu incentrata sull’azione di Dermestes, coleottero che nelle successioni classiche è normalmente collocato solo nelle fasi tardive. La sua massiccia presenza su corpi deceduti da poco tempo trova spiegazione nella rapidità in cui i corpi in ambiente desertico superano velocemente la fase gassosa e colliquativa, arrivando velocemente alle fasi di essiccamento e mummificazione, nicchia tipica dei Dermestidi. Si può supporre che Dermestes sia giunto sui corpi dopo qualche giorno dal decesso a causa delle condizioni geografiche (il deserto può essere ostile anche per un insetto necrofago) e ambientali (l’incidente aereo era avvenuto a febbraio) che hanno rallentato anche i tempi di copula e di riproduzione. L’autore dello studio ritenne improbabile la possibilità che una sola generazione di coleotteri aveva potuto scheletrizzare il corpo di un uomo adulto a quelle temperature, quindi suppose una doppia generazione per un totale di circa novanta giorni dalla data in cui i resti furono rinvenuti. Il dato fu poi supportato dalle notizie riguardanti l’incidente aereo, avvenuto 100 giorni prima del ritrovamento dei corpi (Bellussi, 1933).
6) Correlazioni tra entomofauna e scena del crimine.
Caso 1. Il mese di gennaio venne rivenuto nella città di Panguipulli (Cile) il corpo di una donna di 60 anni, all’interno del suo appartamento. Il cadavere si trovava sul letto, coperto da uno strato di terriccio e presentava lesioni da punta e taglio. La polizia indicò come sospettato il figlio ventiduenne, che negò ogni responsabilità circa la morte della madre. Durante l’ispezione nell’appartamento del giovane non furono trovate tracce di sangue della vittima, fu però portato in laboratorio un paio di scarpe da ginnastica con la suola sporca di detrito simile a quello che copriva il cadavere. La comparazione non diede informazioni utili. Ma nelle scanalature delle scarpe vennero repertati dei pupari, identificati come appartenenti a Calliphora sp. uguali a quelli presenti nel terriccio della scena del crimine in cui erano visibili anche resti di adulti. Fu possibile concludere che il sospettato era stato nel luogo in cui erano presenti i pupari, quindi dove si trovava il corpo in decomposizione (Tosti-Croce and Leyton, 2006).
Caso 2. Il corpo nudo di una donna venne trovato poco lontano da una strada, nei sobborghi di una città della California. Durante la mattinata un sergente del Dipartimento dello Sceriffo condusse un sopralluogo preliminare e la mattina seguente notò delle macchie rosse e pruriginose sulla sua pelle. Egli interpretò queste come morsi delle larve dell’acaro del tutto simili a quelle subite anni prima quando si trovava in Kentucky. In seguito venne a sapere che anche 20 dei 23 membri della squadra avevano lesioni simili e ragionò sul fatto che fosse inusuale la presenza di quel particolare acaro in California. Nell’osservare le fotografie dei sospettati dell’omicidio egli notò una persona con le stesse macchie rosse e concluse che doveva esserci una correlazione tra i morsi dell’acaro, il sospettato, la squadra di soccorso e la scena del crimine. Venne interpellato un entomologo e portato sul luogo di ritrovamento del corpo, furono riesaminate le fotografie e venne fatto un sopralluogo nella casa in cui il sospettato aveva affermato di aver passato la notte dell’omicidio. L’indagine sull’area della scena di morte portò alla raccolta di un gran numero di larve di acaro, specialmente nella radura dove era stato rinvenuto il cadavere, mentre le aree vicine con habitat simili o diversi resero un numero ridottissimo di campioni repertati, probabilmente a causa della predazione da parte di lucertole.
È stato quindi possibile constatare che la probabilità di incontrare un numero di esemplari tali da provocare lesioni a più di 20 persone, in una zona diversa da quella del ritrovamento del corpo, era remota. Questa osservazione, insieme all’analisi delle caratteristiche dei morsi sulla pelle dei membri della squadra e del sospettato furono presentate al processo, alla fine del quale l’imputato fu condannato (Catts & Haskell, 1990).
7) Influenza degli insetti sul Bloodstain Pattern Analysis (B.P.A.)
Nel 1997 due uomini venero trovati uccisi da colpi di arma da fuoco alla testa ed al torace in un appartamento del Nebraska. Numerose aree interessate da macchie di sangue (ing. Bloodstain) da bassa, media e alta velocità furono osservate sia nelle immediate vicinanze dei cadaveri che su alcuni suppellettili e sulle pareti più lontane della stanza; furono inoltre rinvenute simili aree sulle porte e sulle pareti di altre stanze della casa. Gli investigatori ritennero in un primo momento che l’omicidio fosse stato preceduto da una violenta colluttazione e/o che i corpi fossero stati spostati dal teatro dell’uccisione. La spiegazione poteva quindi essere ipotizzata come una reazione delle vittime ad un tentativo di rapina (o ad un litigio finito in tragedia) piuttosto che ad un’esecuzione. Ciononostante nessun segno di lotta era presente nella casa e nella stanza del ritrovamento. Un’attenta analisi delle macchie di sangue presenti su tutte le aree della casa permise di distinguere quelle originate dall’evento omicidiario da quelle che non presentavano i parametri classici degli “sputters” (trad.= spruzzi) da ferita da arma da fuoco. Quest’ultime in particolare presentavano caratteristiche simili: diametro compreso tra 1-3 mm, forma simile ad uno spermatozoo, assenza di direzionalità univoca nelle aree interessate (fig.24).

A seguito dell’intervento di un entomologo, si provò a riprodurre tale fenomeno ponendo delle mosche (della stessa specie di quelle rinvenute sulla scena del crimine) a contatto con un liquido di colore rosso in una stanza chiusa. Dopo alcuni giorni, il rigurgito ed il contatto delle mosche produsse un effetto assolutamente sovrapponibile a quello della scena criminis. Le macchie sparse in tutta la casa furono quindi attribuite alle mosche, il che indirizzò le indagini verso l’ipotesi di un’esecuzione premeditata da parte di chi aveva commesso il reato (Benecke et al., 2003).
Questo caso dimostra come la conoscenza dei fenomeni indotti dall’entomofauna presente sulla scena del crimine può influenzare significativamente le indagini. Molti specialisti del Blood Pattern Analysis “mettono in guardia” circa le valutazioni errate che possono essere indotte ignorando il contributo degli insetti (James, 1998; Bevel, 1997).
8) Ferite causate dall’entomofauna necrobionte.
Il corpo completamente vestito di una giovane donna fu trovato in un terreno con poca vegetazione in Maryland (USA). I resti presentavano grandi masse di larve sul petto e sul collo e altre infestazioni si notavano sui palmi delle mani. Gli insetti non furono campionati, ma furono scattate numerose fotografie del corpo e del luogo di rinvenimento. Le foto furono sottoposte all’esame degli entomologi perché stabilissero se l’età delle larve fosse coerente con la data della presunta scomparsa della vittima. Ma, analizzando il materiale fotografico gli esperti poterono indicare la presenza di traumi al momento della morte non riconducibili, come descritto inizialmente, all’attività degli artropodi. Quando questa informazione fu esaminata dalle autorità, la vittima era già stata sepolta e la morte classificata come “decesso da overdose”. Sulla base delle prove entomologiche la magistratura ordinò la riesumazione ed una nuova autopsia che infine rilevò incontrovertibili segni di ferite da coltello su cui le larve avevano, dopo la morte, compiuto i loro primi stadi vitali (Catts & Haskell, 1990).
9) Entomotossicologia.
Caso 1. Il corpo non identificato di una donna fu rinvenuto in una zona rurale della Finlandia. Il cadavere era in avanzato stadio di decomposizione, colonizzato da larve di ditteri, che vennero prelevati e messi in camera di crescita; in seguito venne analizzato il contenuto di mercurio degli insetti adulti. La bassa concentrazione di mercurio contenuta negli insetti adulti indicava che la vittima proveniva da una zona poco inquinata, a differenza del luogo di ritrovamento del corpo. Ulteriori analisi permisero in seguito di accertare questa ipotesi (Nortueva, 1977).
Caso 2. Il corpo senza vita di una donna di 22 anni fu trovato da un escursionista in un’area boscosa, vicino al letto di un torrente. Il cadavere era completamente scheletrizzato, coperto dagli indumenti intatti ad eccezione delle lacerazioni dovute alle intemperie del periodo post-mortem. Vicino al corpo erano presenti una lettera ed una borsa contenente i documenti, una scatola di medicinali vuoti e una prescrizione medica di Phenorbital.
Dopo l’identificazione, eseguita anche con esami dentali, si scoprì che la giovane aveva tentato il suicidio già cinque volte. Durante l’autopsia vennero prelevate le larve di insetti per stabilire il P.M.I., ma in mancanza di tessuti per analisi tossicologiche vennero utilizzati gli stessi campioni, che risultarono positivi al Phenorbital.
Gli entomologi consultati identificarono le larve come di Cochliomyia macellaria e assicurarono che la droga presente negli insetti non poteva aver avuto origine da meccanismi metabolici ma dal pabulum di questi (Beyer et al., 1980). In questo caso l’analisi entomotossicologica riuscì a fugare ogni dubbio circa il farmaco utilizzato per il suicidio anche nell’impossibilità di perseguire le tecniche ortodosse della tossicologia forense.
10) Selezione dell’entomofauna dovuta allo spostamento, al seppellimento, all’immersione ed all’occultamento.
Caso 1. Il corpo di una donna fu scoperto in un boschetto ombroso adagiato sul terreno, con la testa contusa, le mani legate all’altezza dei polsi e le caviglie strette con una fune.
Le larve più grandi (di Cynomya cadaverina) si trovavano soprattutto sul volto della vittima e indicavano uno sviluppo di circa 5 giorni ma questo insetto colonizza i cadaveri solo quando lo stadio di decomposizione fresca è avanzato; quindi l’ipotesi degli entomologi fu che il crimine venne commesso qualche giorno prima in un luogo diverso da quello del ritrovamento e che Cynomya cadaverina giunse solo dopo sul corpo. Altre prove della scena di morte erano in accordo con questa conclusione (Catts & Haskell, 1990).
Caso 2. Il corpo di una donna anziana venne rinvenuto nel mese di marzo sul pavimento di un alloggio della città di Venezia. Il cadavere, coperto da un vestito di lana, si trovava completamente coperto da materiale di varia natura tra cui anche cuscini e materassi. Durante l’autopsia, che certificò la morte causata da un attacco di cuore, furono campionati ditteri delle specie Calliphora vicina, Fuscina stabulans e del genere Megaselia. La strana preponderanza di esemplari di Megaselia a scapito dei Calliforidi, avrebbe potuto falsare le considerazioni entomo-tanatologiche. In realtà l’evento fu brillantemente spiegato osservando che gli adulti di Megaselia depongono le uova su piante e animali in decomposizione e nel suolo da cui le larve si muovono scavando anche a profondità di 50 cm. Questo comportamento può spiegare il motivo della loro massiccia presenza: le larve, attratte dal corpo in decomposizione, erano state in grado di “scavare” attraverso le coperture del corpo, mentre solo pochissimi esemplari delle altre specie erano stati in grado di raggiungere il corpo (Turchetto et al., 2001).
Caso 3. In novembre furono rinvenute numerose carcasse di suino, interrate abusivamente, in un terreno privato poco lontano dalla città del nord Italia. Fu aperta un’inchiesta per la valutazione del tempo d’interramento degli animali e vennero chiamati, con l’incarico di periti, esperti nelle discipline di Medicina Veterinaria, Anatomo-Patologia Veterinaria ed Entomologia Forense. Durante il sopralluogo sul terreno incriminato furono scavate, con l’ausilio di una scavatrice meccanica, 6 fosse di circa 2 metri di profondità, in zone scelte arbitrariamente: in 4 dei 6 punti di escavazione furono trovati resti di carcasse di suino in diversi stadi di decomposizione. Nella prima fossa si trovarono resti di carcasse, molte larve e pupe di Ditteri (Helina sp. e Calliphora vicina), alcuni adulti e larve di Coleotteri, mentre nelle altre fosse si evidenziarono solo carcasse od ossa di suini, nessun reperto entomologico.
Le carcasse rinvenute nella prima buca presentavano alterazioni cadaveriche compatibili con un periodo d’interramento variabile tra 6 mesi e 1 anno, confermato dallo studio entomologico delle pupe di Helina sp.; tale dittero è in grado di colonizzare carcasse sepolte deponendo uova sul terreno da cui schiudono larve capaci di scendere verso il corpo sepolto. Inoltre, la presenza di altre larve di Helina sp., databili come deposte da 1 mese, permise di ipotizzare la sovrapposizione delle carcasse in due tempi successivi d’interramento (Appino et al., 2005). I campioni di Calliphora vicina, specie di superficie, permisero invece di accertare che alcune carcasse erano state precedentemente esposte alla superficie e solo in un secondo momento inumate. I reperti delle altre fosse furono datati un tempo massimo di 2 anni, periodo confermato dall’assenza di reperti entomologici, deteriorati nel terreno. L’insieme di tali risultati permise di accertare il compimento ripetuto e continuato di una condotta illecita.
Caso 4. Durante un’immersione ricreativa due subacquei trovarono un’automobile, capovolta sul fondo del fiume Muskegon (Michigan). All’interno fu rinvenuto il corpo di una donna. La polizia estrasse la macchina dal fiume e risultò essere di proprietà del marito della vittima. Sul capo della donna i medici rilevarono contusioni che sembravano non essere state procurate dall’incidente. Il marito dichiarò di non aver più visto la donna dalla sera nebbiosa di settembre in cui si era consumato un litigio, al seguito del quale la moglie era andata via in macchina. L’automobile sembrava in buone condizioni: ciò faceva presupporre che fosse stata spinta, piuttosto che essere precipitata nel fiume in seguito ad un incidente. Sebbene l’acqua fredda avesse conservato il corpo, il P.M.I. era in ogni modo di difficile determinazione. Sull’auto gli investigatori trovarono e campionarono diversi insetti acquatici attaccati al parabrezza, al paraurti e alle portiere. Tra gli altri artropodi acquatici vennero anche rinvenuti pupari di Simulidi. Le femmine adulte dei Simulidi depongono le uova tra la fine della primavera e l’inizio dell’estate. Queste rimangono nel sedimento del fondale e vanno obbligatoriamente in diapausa fino all’autunno o l’inverno, dopodiché si schiudono in larve che si attaccano ad un substrato (rocce o vegetazione, in questo caso un’automobile) e si nutrono per filtrazione del particolato presente in acqua. Dopo poco, tra marzo e aprile le larve entrano in “pupazione” ed in seguito (intorno alla metà di maggio) emergono insetti adulti che escono dall’acqua e continuano il ciclo vitale come insetti alati. Basandosi sull’identificazione della specie e sulla conoscenza del ciclo di vita dei Simulidi presenti sulla macchina è stato possibile dichiarare che l’automobile doveva essere precipitata in acqua presumibilmente in autunno. Il sospettato aveva in precedenza dichiarato di aver avuto notizie dalla moglie durante l’inverno e la primavera ma ciò non era possibile in base ad alcuni parametri autoptici ed ai reperti entomologici.
Grazie alle informazioni ottenute dall’autopsia, dall’analisi degli insetti e basandosi su altre prove presentate dall’accusa e dai testimoni, fu provato che la donna era scomparsa l’autunno precedente e che la macchina si trovava nel fiume da circa nove mesi; ciò fu ritenuto sufficiente per condannare il marito per omicidio di secondo grado (Byrd & Castner, 2001).
11) Assenza di entomofauna.
Il corpo di un uomo venne ritrovato disteso prono sul pavimento della toilette di una stazione di servizio. Sul cadavere non vi erano evidenze di ferite, ma un braccio era stretto da laccio emostatico e poco lontano dal corpo era presente una siringa. L’uomo era sparito da 17 giorni e la sua automobile, ritenuta abbandonata nel parcheggio della stazione di servizio, era stata rimossa giorni prima del ritrovamento del corpo. Gli inservienti dei servizi igienici, non potendo entrare nella toilette a causa della porta chiusa dall’interno, non si erano accorti della presenza del cadavere. Soltanto molti giorni dopo l’odore era diventato talmente forte da insospettire i gestori al punto di prendere la decisione di abbattere la porta. In sede di autopsia non fu possibile rinvenire campioni entomologici.
La spiegazione più plausibile della mancanza di insetti si trova nel potente sistema di aerazione che eliminava gli odori dai bagni della stazione di servizio e nell’ampia grata disposta sul pavimento che ha aiutato nella percolazione dei liquami putrefattivi. L’assenza di insetti sulla scena di morte è possibile se le temperature sono molto alte o molto basse e/o in condizioni particolari di mantenimento del corpo, come luoghi sigillati, inumazione, immersione e congelamento. In condizioni normali, la totale assenza di insetti per tutta la durata della decomposizione è una possibilità remota, perché questi animali hanno caratteristiche fisiologiche che permettono loro il ritrovamento di un corpo in decomposizione anche a grande distanza e caratteristiche morfologiche che gli consentono l’invasione di quasi ogni luogo. Quindi in mancanza delle succitate condizioni inibenti lo sviluppo dell’entomofauna diventa importante non sottovalutare che sulla scena del delitto “l’assenza di tracce è di per se una traccia”, principio che vale per tutte le applicazioni della criminalistica. Una volta eliminate tutte le possibili cause di isolamento e/o inibizione dello sviluppo è dunque verosimile ipotizzare uno “staging” attuato tramite alterazioni e spostamenti del cadavere.
12) Maltrattamenti e Negligenze.
Caso 1. In un appartamento che versava in cattive condizioni di ordine e pulizia, alle 6.35 del mattino venne rinvenuto il corpo senza vita di una donna di 89 anni, che abitava insieme alla figlia. Il cadavere fu rimosso senza l’intervento di un medico e dall’osservazione delle condizioni del corpo gli inquirenti datarono la morte a molti giorni prima.
Venne aperto un procedimento per occultamento di cadavere nei confronti della figlia che però affermava di aver servito la cena alla madre la sera prima del ritrovamento. L’autorità giudiziaria ordinò un esame del corpo da parte di un anatomo-patologo il quale campionò larve di ditteri al secondo e terzo stadio. In assenza di altre informazioni tali esemplari avrebbero rappresentato il segnale di un certo lasso temporale trascorso dalla morte. Invece l’esame esterno rese evidenti estese piaghe da decubito sulla schiena e sulle gambe, erroneamente interpretate dagli inquirenti come segni di putrefazione. L’analisi medico-legale congiuntamente a quella entomologica permisero infine di accertare un’infestazione da parte degli insetti iniziata alcuni giorni prima del decesso, avvenuto in realtà nelle ore precedenti al ritrovamento del corpo. Caddero quindi i requisiti per un procedimento penale per occultamento di cadavere, ma fu valutata la condotta negligente della figlia dei confronti dell’anziana madre (Gherardi et al., 2004).
Caso 2. Un caso molto discusso a livello mediatico riguardò un uomo giunto all’ospedale di Chicago ferito alla gamba a causa di un incidente. Dopo diverse operazioni l’uomo fu dimesso ripresentandosi, però dieci giorni dopo al pronto soccorso dell’ospedale con larve che fuoriuscivano dalla ferita della gamba. Gli avvocati d’entrambe le parti interrogarono i propri esperti per risalire alla data in cui tale infestazione si era originata e quindi per stabilire eventuali responsabilità. Gli entomologi stabilirono che l’infestazione era attribuibile a Phaenicia sericata, mosca abbondante nella zona di Chicago in quel periodo dell’anno. Assumendo che la temperatura sotto le bende fosse compresa tra i 29°C e i 35°C si poté stabilire che, in tali condizioni, il tempo necessario a Phaenicia sericata per raggiungere lo stadio post-feeding è quantificabile in tre giorni dalla deposizione. In quella data il paziente si trovava in casa e non nell’ospedale che fu scagionato da ogni responsabilità (Greenberg, 1984).
8. Conclusioni e prospettive future
Come evidenziato nei paragrafi precedenti, la precisa stima del P.M.I. rimane il dato più indicativo dell’approccio entomologico alle indagini medico-legali. Molti aspetti però devono ancora essere approfonditi, a causa delle innumerevoli variabili esistenti, per dare piena dignità probatoria ad uno strumento investigativo divenuto oramai irrinunciabile. Anzitutto formare le coscienze e le conoscenze del personale che normalmente interviene sulla scena del crimine: annotare e documentare con precisione certosina le condizioni ambientali in rapporto agli stadi di decomposizione del corpo ed all’influenza reciproca che entrambi possono esercitare sulla biocenosi cadaverica.
La conoscenza dell’etologia e della fisiologia degli insetti è inoltre ancora incompleta e contribuisce ad allargare uno spettro delle variabili: la lunghezza del giorno (fotoperiodo), le temperature ambientali e cadaveriche, la tendenza diurna o notturna all’ovodeposizione, i fenomeni di diapausa, competizione, parassitismo e predazione alterano drasticamente lo sviluppo dell’entomofauna (Amendt et al., 2004). Ma se a ciò si aggiungono situazioni d’isolamento fisico, occultamento (es. tramite materiali sigillanti), spostamento, esposizione ad agenti chimici e fisici (es. corpi carbonizzati), presenza di fauna non specializzata (formiche, ratti, altri mammiferi, etc.), seppellimento ed immersione dei cadaveri si comprende come l’interpretazione delle variabili diventi estremamente complessa. Il ruolo dell’acqua nei fenomeni degradativi cadaverici e nella colonizzazione degli artropodi acquatici ha ricevuto la minor attenzione da parte degli studiosi, non esistendo ad oggi dei modelli affidabili (Amendt et al. 2004). Lo studio delle conoscenze scientifiche si sta, nell’ultimo decennio, sempre più consolidando così come l’esperienza derivante dall’applicazione a casi reali.
Per ciò che attiene all’approccio biomolecolare all’Entomologia Forense, il maggiore ostacolo consiste nella mancanza di dati genotipici dell’entomofauna italiana. Le banche dati genetiche per le specie d’interesse forense sono per lo più frutto di studi condotti su popolazioni di insetti nelle aree nordeuropee ed americane e non possono essere applicate indiscriminatamente sugli esemplari campionati sul nostro territorio. Considerata l’elevata variabilità genetica ed ambientale in campo entomologico, non sarebbe rigoroso estrapolare i dati in conformità a comparazioni che non tengono conto della tipicità del nostro ecosistema. D’altronde le valutazioni statistiche che ponderano l’unicità dei profili genotipici umani, ordinariamente utilizzate nelle nostre aule di Giustizia, sono considerate attendibili grazie ai confronti con i consolidati dati genotipici riferiti ai differenti ceppi della popolazione mondiale.
è quindi auspicabile, da parte dei nostri atenei, il potenziamento della ricerca di base in tale settore. Moltissime speranze sono poi riposte nello sviluppo dei marcatori molecolari “RNA-based” (analisi dei trascritti mediante mRNA differential display) e proteomici (fattori della trascrizione tra cui TATA-box binding protein) per l’identificazione univoca dello stadio di sviluppo dell’insetto (Ames et al., 2004). Ciò permetterebbe di velocizzare e semplificare notevolmente le attuali attività di laboratorio producendo un risultato difficilmente “attaccabile” nel contenzioso penale. Infine è ipotizzabile che tra qualche tempo saranno sviluppati marcatori molecolari in grado di identificare, come per l’uomo, il singolo insetto, permettendo la tracciabilità di un dato esemplare lungo tutto i suoi cicli vitali.
La correlazione tra il cadavere e l’entomofauna mediante l’analisi del DNA umano presente all’interno degli artropodi richiede la predisposizione del laboratorio interessato per l’applicazione di metodiche estrattive “high-purity” e per le analisi in alta sensibilità (DNA-LCN). Inoltre, come anche già evidenziato, un ulteriore limite è anche offerto dalla scarsa attenzione/conoscenza prestata a quest’approccio da parte di chi opera nel sopralluogo e dei medici legali in sede di autopsia (Gomes et al., 2006). In un futuro prossimo l’analisi genotipica trarrà comunque notevole vantaggio dallo sviluppo della tecnologia dei microarray che permetterà, tra le altre cose, anche la tipizzazione del genotipo umano e dell’insetto con una singola analisi.
In conclusione l’entomologia forense rappresenta uno strumento investigativo e giudiziario indispensabile unitamente a tutte le altre branche della criminalistica. Lo sforzo umano ed economico impiegato da molti Paesi viene costantemente ripagato dalla risoluzione di casi giudiziari soprattutto di quelli definiti “senza apparente motivo”. In Italia tale studio è attualmente ad appannaggio di pochi ricercatori che con estrema dedizione e elevata professionalità trainano la ricerca in questo affascinante mondo degli insetti: a questi le forze di polizia si stanno affiancando soprattutto negli ultimi anni, come le recenti cronache testimoniano. Grazie alle collaborazioni con scienziati nel precipuo settore, infatti, si cerca di diffondere la cultura dell’entomologia nonché, soprattutto, l’immediato risvolto pratico. Le aule dibattimentali, notoriamente prudenti verso le “nuove esperienze”, si stanno aprendo a questo nuovo mondo, contribuendo, con la casistica giuridica, a spingere verso la necessità di impiegare le larve come “nuove fonti di prova”.
Un autorevole scienziato forense diceva che “su una scena del crimine ogni “evidence” parla: sta alla capacità dell’operatore saper ascoltare” ed il mondo degli insetti sicuramente rappresenta un coro che, almeno al momento non sempre siamo in grado di apprezzare.
“Era stata finora occulta la generazione di questo terribile insetto, ed a me è toccata la sorte di scoprirla” (A. Vallisneri, 1733).
___________
(*) - Ufficiale addetto al Reparto Carabinieri Investigazioni Scientifiche di Messina.
(**) - Responsabile del Forensic Entomology Laboratory (F.E.Lab) - ASL 1 Torino.
(***) - Comandante la Sezione di Biologia Molecolare del Reparto Carabinieri Investigazioni Scientifiche di Messina.
(1) - Dal latino Insecare = tagliare, con questo termine si intende indicare gli animali il cui corpo è “tagliato” o suddiviso in pezzi.
(2) - Dal greco Entèmnein = tagliare in pezzi, e dal latino forenseum = attinente al “foro”.
(3) - dal latino Fàunus, antico dio protettore degli armenti e delle greggi.
(4) - Dal greco Sapros = decomposto e Phagein = mangiare.
(5) - Dal greco Nekros = morto, e Biotos = vitale, quindi “che vive su ciò che è morto”.
(6) - Dal greco Nekros = morto e Phagein = mangiare.
(7) - Insieme di tutti gli organismi.
(8) - Dal greco Dis (due), Pteron (ala).
(9) - Dal greco Koleos (astuccio) e Pteron (ala).
(10) - La quantità di calore richiesta per lo sviluppo di un insetto dall’ovodeposizione ad un qualsiasi stadio di sviluppo può essere calcolata in termini di gradi/ore accumulati (ADH, Accumulated Degree Hours) o di gradi/giorni accumulati (ADD, Accumulated Degree Days, giorni): ogni stadio di sviluppo richiede una precisa “quantità di temperatura” e ogni specie ha bisogno di un numero specifico di gradi/ora o gradi/giorno per completare il suo sviluppo a temperature definite.
(11) - Successo iniziale o Fallimento finale.
(12) - Sia per le indagini di P.G. sia nell’identificazione di cadaveri in casi di mass fatality.
(13) - Low copy number: DNA in basso numero di copie.
(14) - Una sorta di “contrassegni” molecolari.
(15) - Tassonomia, dal greco taxinomia, che deriva dalle parole taxis = ordine, e nomos = regole.